

Queuine links translational control in eukaryotes to a micronutrient from bacteria
- PMID: 30715423
- PMCID: PMC6468285
- DOI: 10.1093/nar/gkz063
Queuine links translational control in eukaryotes to a micronutrient from bacteria
Abstract
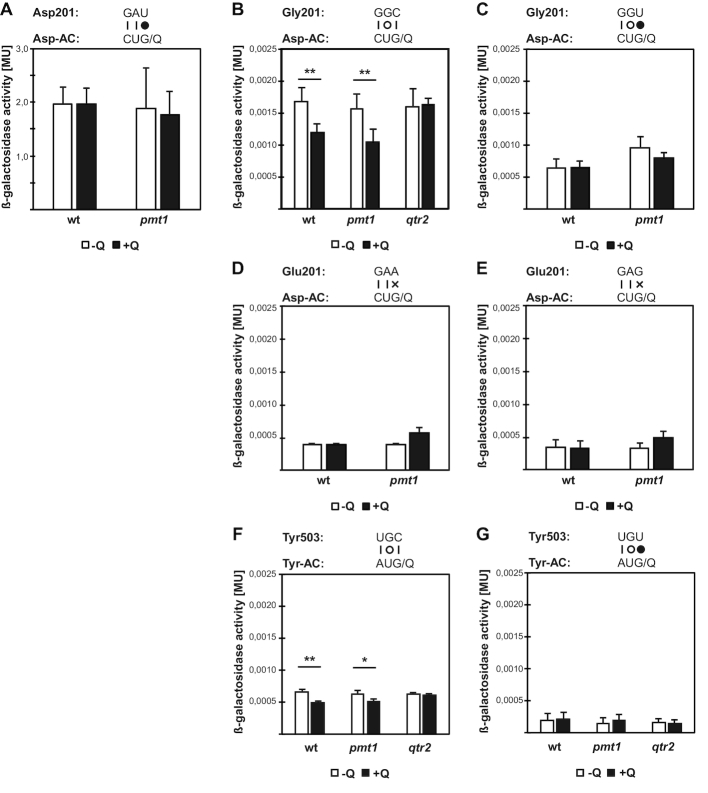
In eukaryotes, the wobble position of tRNA with a GUN anticodon is modified to the 7-deaza-guanosine derivative queuosine (Q34), but the original source of Q is bacterial, since Q is synthesized by eubacteria and salvaged by eukaryotes for incorporation into tRNA. Q34 modification stimulates Dnmt2/Pmt1-dependent C38 methylation (m5C38) in the tRNAAsp anticodon loop in Schizosaccharomyces pombe. Here, we show by ribosome profiling in S. pombe that Q modification enhances the translational speed of the C-ending codons for aspartate (GAC) and histidine (CAC) and reduces that of U-ending codons for asparagine (AAU) and tyrosine (UAU), thus equilibrating the genome-wide translation of synonymous Q codons. Furthermore, Q prevents translation errors by suppressing second-position misreading of the glycine codon GGC, but not of wobble misreading. The absence of Q causes reduced translation of mRNAs involved in mitochondrial functions, and accordingly, lack of Q modification causes a mitochondrial defect in S. pombe. We also show that Q-dependent stimulation of Dnmt2 is conserved in mice. Our findings reveal a direct mechanism for the regulation of translational speed and fidelity in eukaryotes by a nutrient originating from bacteria.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures






References
-
- Ogle J.M., Carter A.P., Ramakrishnan V.. Insights into the decoding mechanism from recent ribosome structures. Trends Biochem. Sci. 2003; 28:259–266. - PubMed
