

TBC1D3 regulates the payload and biological activity of extracellular vesicles that mediate tissue repair
- PMID: 30715917
- PMCID: PMC6463925
- DOI: 10.1096/fj.201802388R
TBC1D3 regulates the payload and biological activity of extracellular vesicles that mediate tissue repair
Abstract
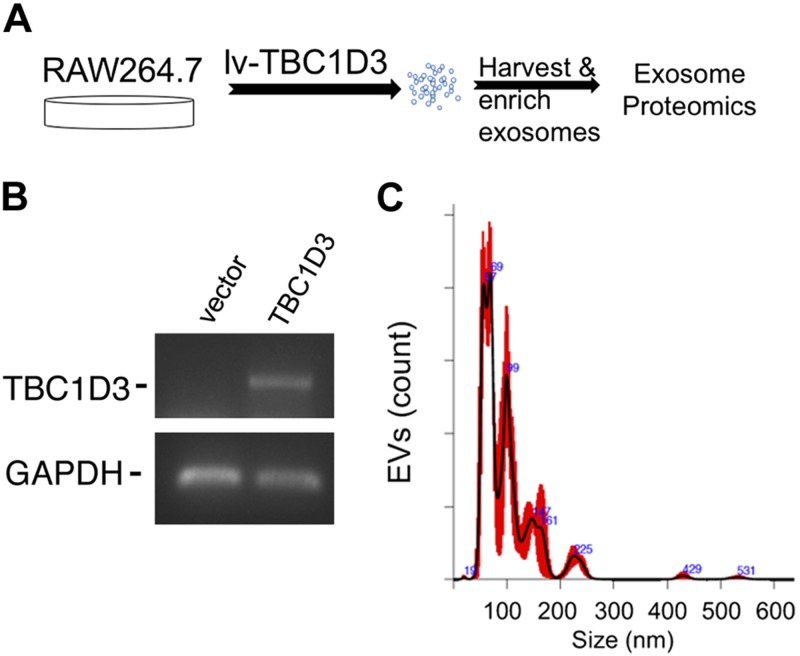
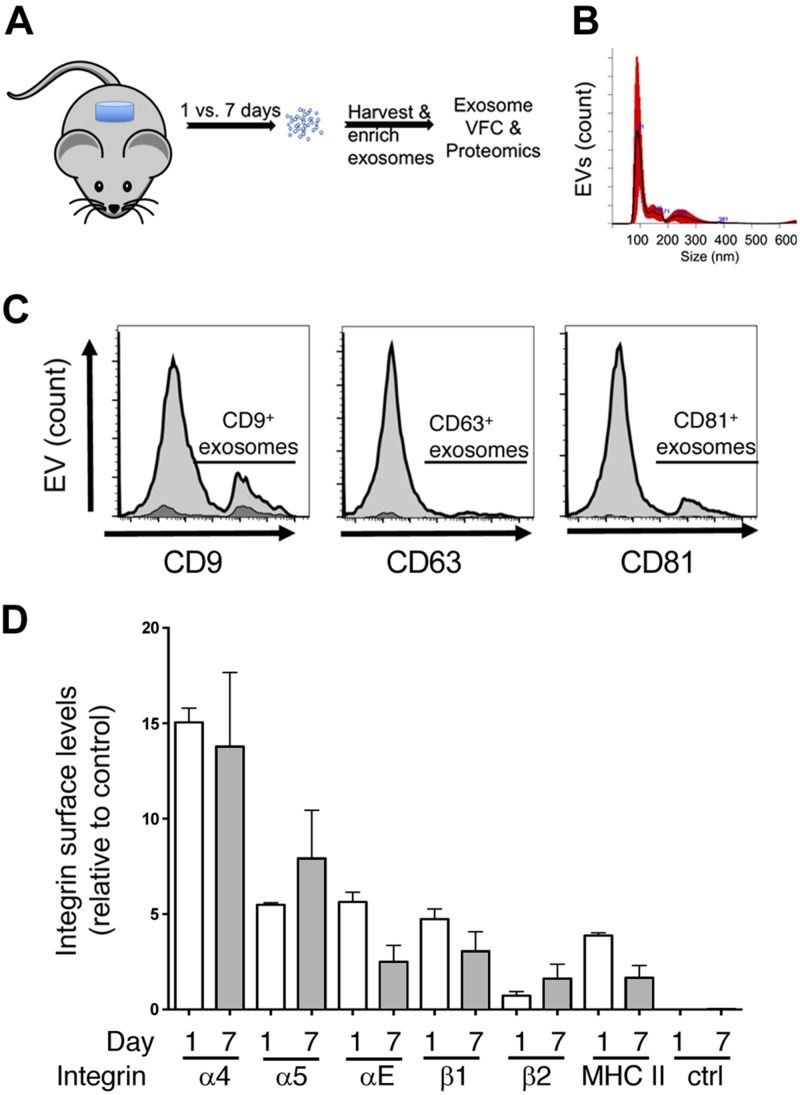
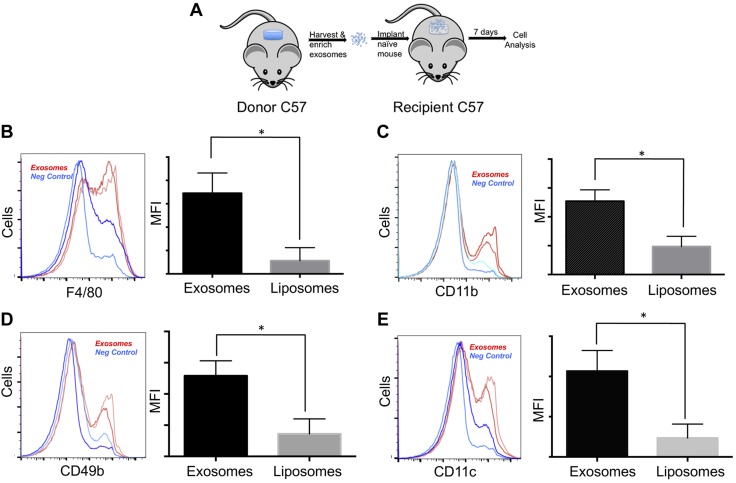
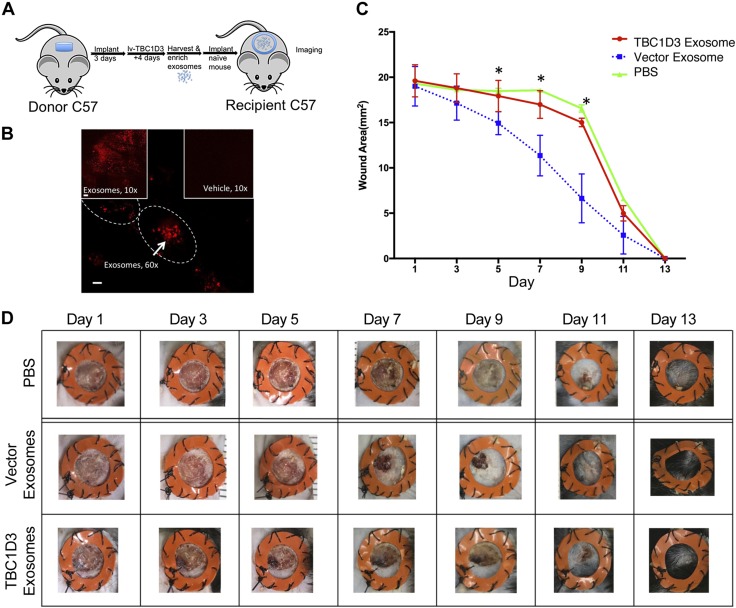
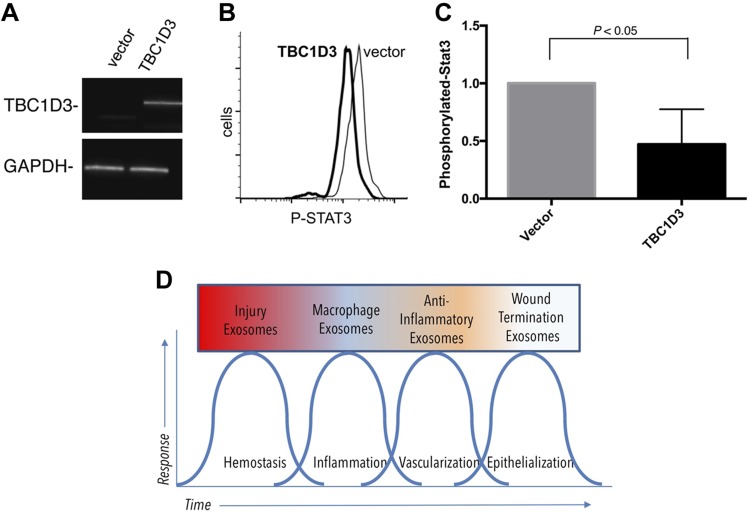
Healthy repair of cutaneous injury is a coordinated response of inflammatory cells, secreted factors, and biologically active extracellular vesicles (EVs). Although constitutive release of EVs into biologic fluids is a hallmark of cultured cells and tumors, their payload and biologic activity appears to be tightly regulated. We show that Tre-2/Bub2/Cdc16 (TBC1) domain family member 3 (TBC1D3) drives the release of an EV population that causes a decrease in phosphorylation of the transcription factor signal transducer and activator of transcription 3 in naive recipient cells. To explore the biologic activity of EVs in vivo, we used a mouse model of sterile subcutaneous inflammation to determine the payload and biologic activity of EVs released into the microenvironment by committed myeloid lineages and stroma. Expression of TBC1D3 in macrophages altered the payload of their released EVs, including RNA-binding proteins, molecular motors, and proteins regulating secretory pathways. A wound-healing model demonstrated that closure was delayed by EVs released under the control of TBC1D3. We show that modulating the secretory repertoire of a cell regulates EV payload and biologic activity that affects outcomes in tissue repair and establishes a strategy for modifying EVs mediating specific biologic responses.-Qin, S., Dorschner, R. A., Masini, I., Lavoie-Gagne, O., Stahl, P. D., Costantini, T. W., Baird, A., Eliceiri, B. P. TBC1D3 regulates the payload and biological activity of extracellular vesicles that mediate tissue repair.
Keywords: RabGAP; exosome; foreign body response; macrophage activation; wound healing.
Conflict of interest statement
The authors thank Ann-Marie Hageny [University of California–San Diego (UCSD)] for expert technical support, and Dr. Antonio DeMaio (UCSD) for use of the Nanoparticle Tracking Analysis Instrument; Dr. Majid Ghassemian (UCSD Biomolecular and Proteomics Mass Spectrometry Facility) provided expert support and analysis for the proteomics studies; Dr. John Nolan (Scintillon Institute, San Diego, CA, USA) provided expert advice on the design of vesicle flow cytometry assays; Dr. Chen Kong (Washington University) helped with consultation on the molecular biology of TBC1D3; and Dr. Lisa A. Hannan provided expert editorial review of the study. Research was supported by operating grants from the U.S. National Institutes of Health, National Institute of General Medical Sciences (1R01GM121530), the UCSD Academic Senate, the UCSD Department of Surgery Reinvestment Fund, the Shock Society, and the Surgical Infection Society. The authors declare no conflicts of interest.
Figures
References
-
- Kojima M., Gimenes-Junior J. A., Langness S., Morishita K., Lavoie-Gagne O., Eliceiri B., Costantini T. W., Coimbra R. (2017) Exosomes, not protein or lipids, in mesenteric lymph activate inflammation: unlocking the mystery of post-shock multiple organ failure. J. Trauma Acute Care Surg. 82, 42–50 - PubMed
-
- Li D., Liu J., Guo B., Liang C., Dang L., Lu C., He X., Cheung H. Y., Xu L., Lu C., He B., Liu B., Shaikh A. B., Li F., Wang L., Yang Z., Au D. W., Peng S., Zhang Z., Zhang B. T., Pan X., Qian A., Shang P., Xiao L., Jiang B., Wong C. K., Xu J., Bian Z., Liang Z., Guo D. A., Zhu H., Tan W., Lu A., Zhang G. (2016) Osteoclast-derived exosomal miR-214-3p inhibits osteoblastic bone formation. Nat. Commun. 7, 10872 - PMC - PubMed
-
- Garzetti L., Menon R., Finardi A., Bergami A., Sica A., Martino G., Comi G., Verderio C., Farina C., Furlan R. (2014) Activated macrophages release microvesicles containing polarized M1 or M2 mRNAs. J. Leukoc. Biol. 95, 817–825 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
