

A chemoproteomic portrait of the oncometabolite fumarate
- PMID: 30718813
- PMCID: PMC6430658
- DOI: 10.1038/s41589-018-0217-y
A chemoproteomic portrait of the oncometabolite fumarate
Abstract
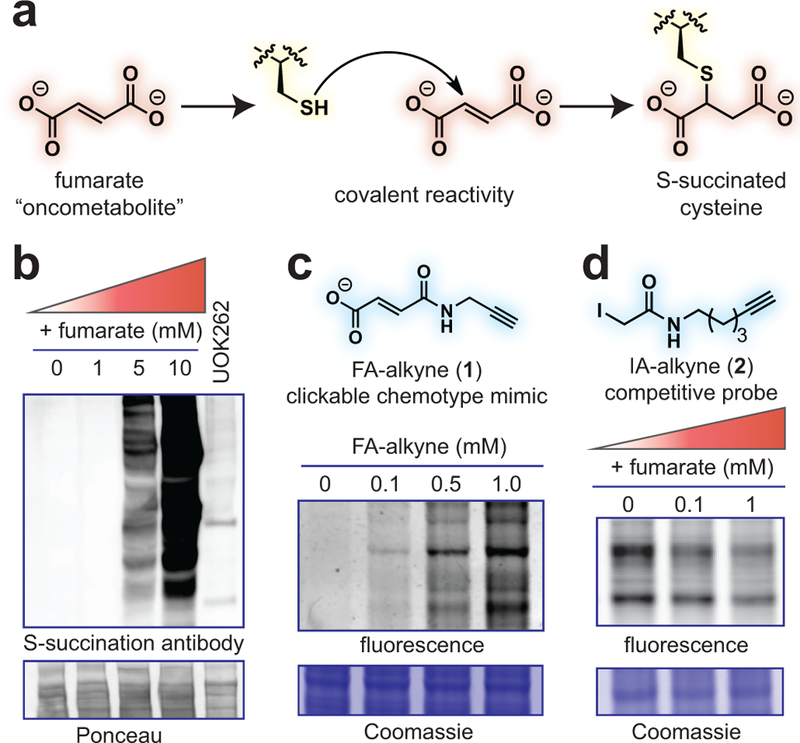
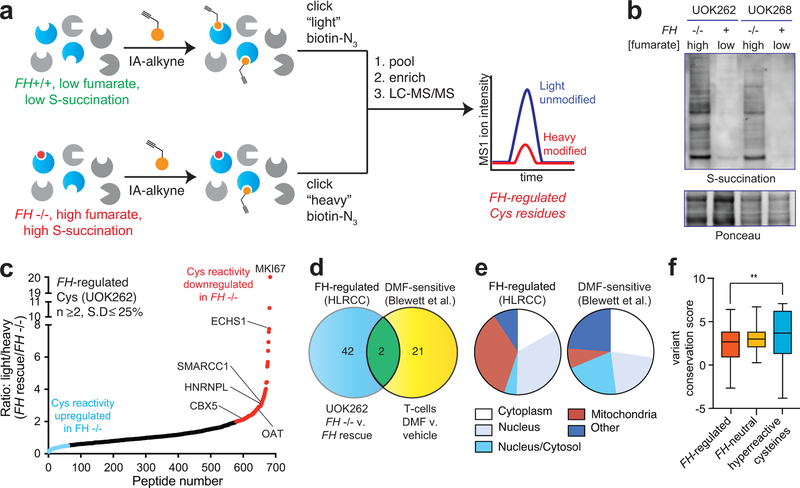
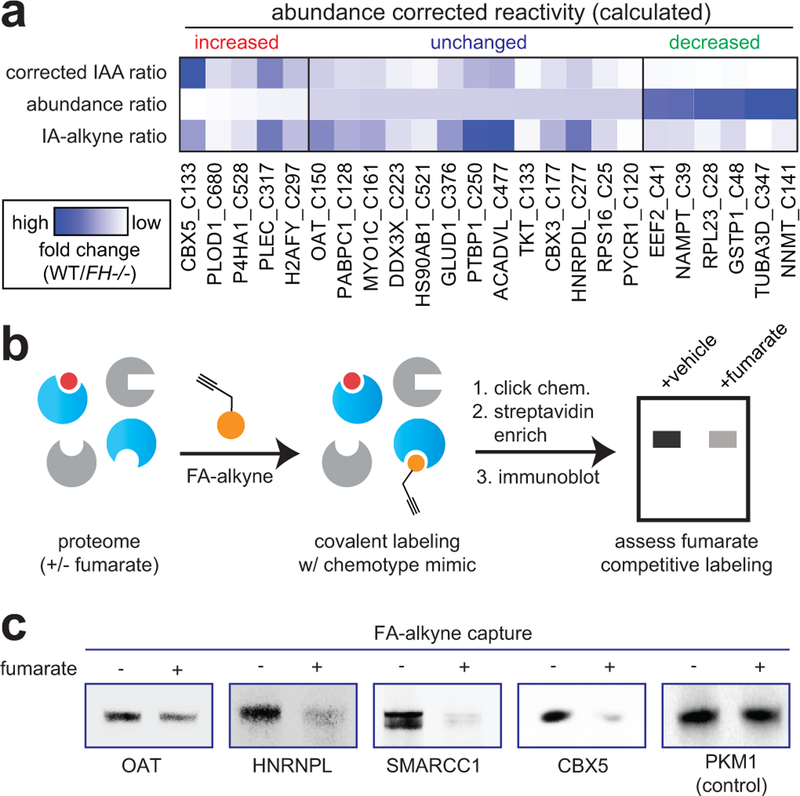
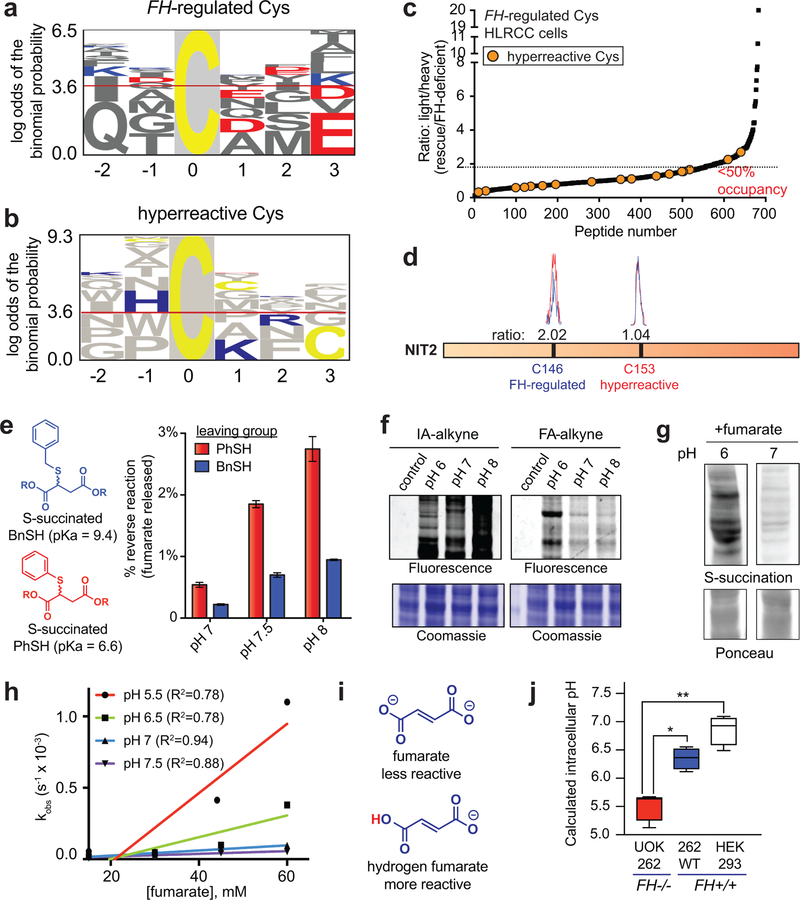
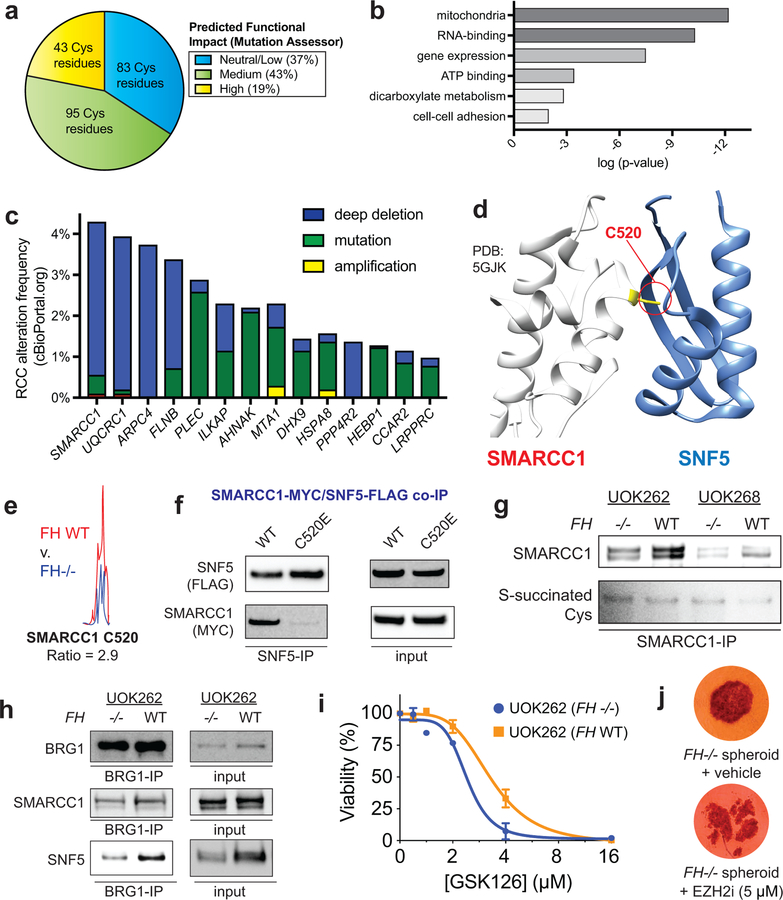
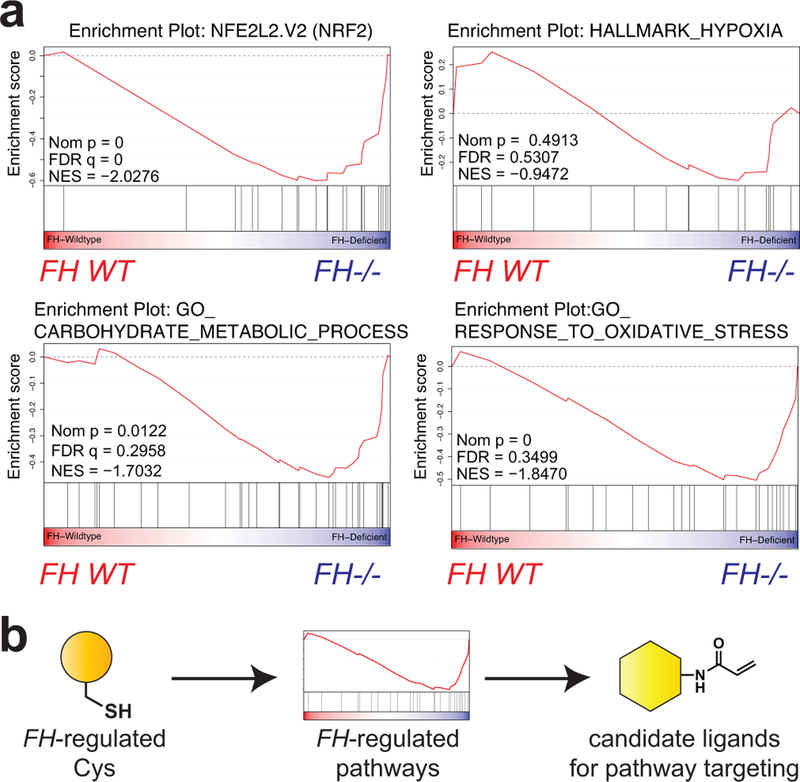
Hereditary cancer disorders often provide an important window into novel mechanisms supporting tumor growth. Understanding these mechanisms thus represents a vital goal. Toward this goal, here we report a chemoproteomic map of fumarate, a covalent oncometabolite whose accumulation marks the genetic cancer syndrome hereditary leiomyomatosis and renal cell carcinoma (HLRCC). We applied a fumarate-competitive chemoproteomic probe in concert with LC-MS/MS to discover new cysteines sensitive to fumarate hydratase (FH) mutation in HLRCC cell models. Analysis of this dataset revealed an unexpected influence of local environment and pH on fumarate reactivity, and enabled the characterization of a novel FH-regulated cysteine residue that lies at a key protein-protein interface in the SWI-SNF tumor-suppressor complex. Our studies provide a powerful resource for understanding the covalent imprint of fumarate on the proteome and lay the foundation for future efforts to exploit this distinct aspect of oncometabolism for cancer diagnosis and therapy.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
