

Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering?
- PMID: 30720746
- PMCID: PMC6387439
- DOI: 10.3390/ijms20030671
Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering?
Abstract
During their lifetime, plants encounter numerous biotic and abiotic stresses with diverse modes of attack. Phytohormones, including salicylic acid (SA), ethylene (ET), jasmonate (JA), abscisic acid (ABA), auxin (AUX), brassinosteroid (BR), gibberellic acid (GA), cytokinin (CK) and the recently identified strigolactones (SLs), orchestrate effective defense responses by activating defense gene expression. Genetic analysis of the model plant Arabidopsis thaliana has advanced our understanding of the function of these hormones. The SA- and ET/JA-mediated signaling pathways were thought to be the backbone of plant immune responses against biotic invaders, whereas ABA, auxin, BR, GA, CK and SL were considered to be involved in the plant immune response through modulating the SA-ET/JA signaling pathways. In general, the SA-mediated defense response plays a central role in local and systemic-acquired resistance (SAR) against biotrophic pathogens, such as Pseudomonas syringae, which colonize between the host cells by producing nutrient-absorbing structures while keeping the host alive. The ET/JA-mediated response contributes to the defense against necrotrophic pathogens, such as Botrytis cinerea, which invade and kill hosts to extract their nutrients. Increasing evidence indicates that the SA- and ET/JA-mediated defense response pathways are mutually antagonistic.
Keywords: hormones; plant defense; signaling pathway.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
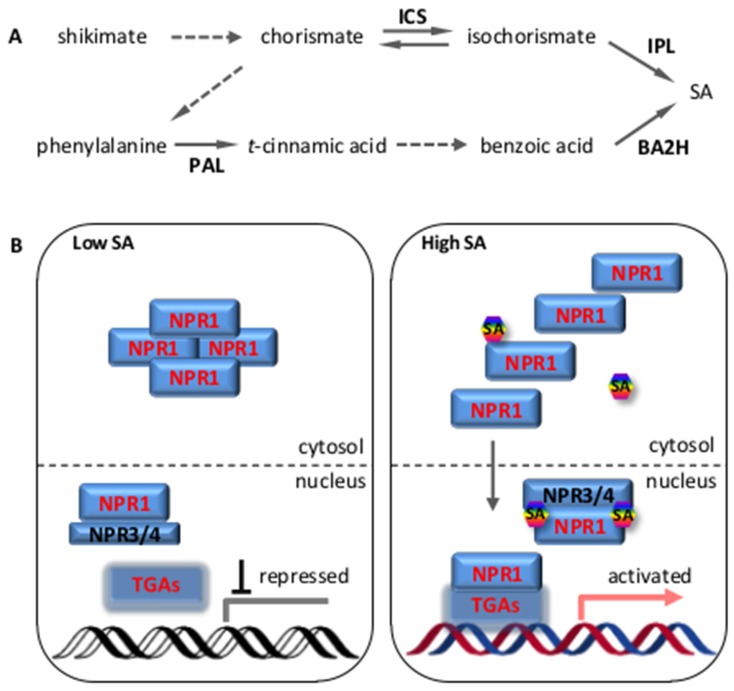
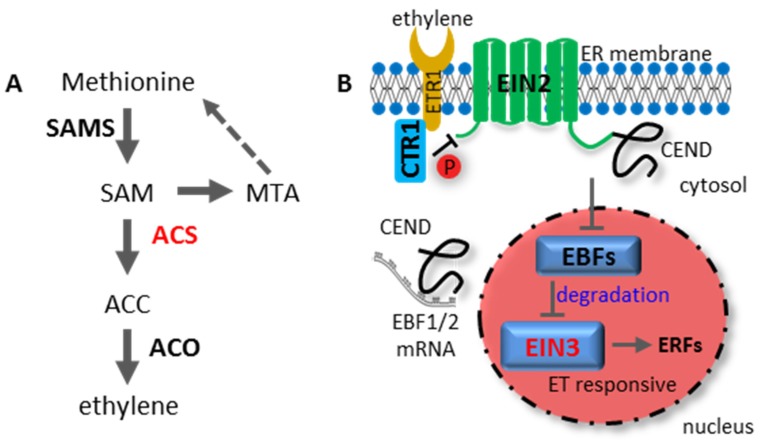
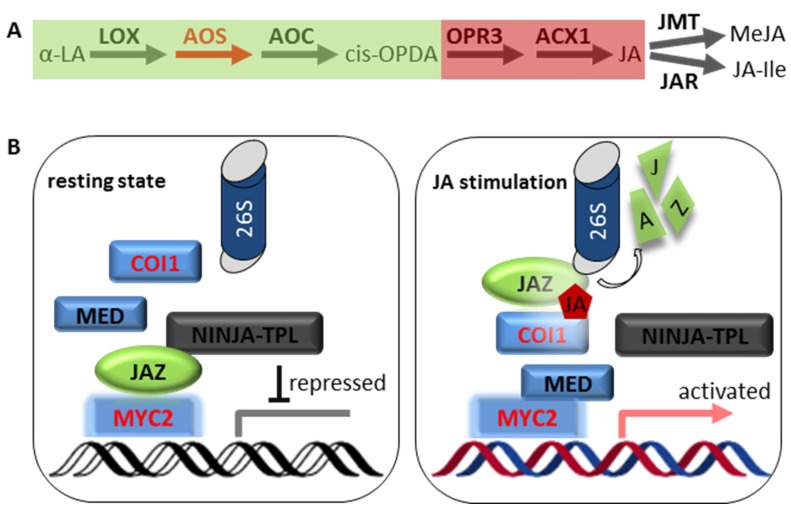
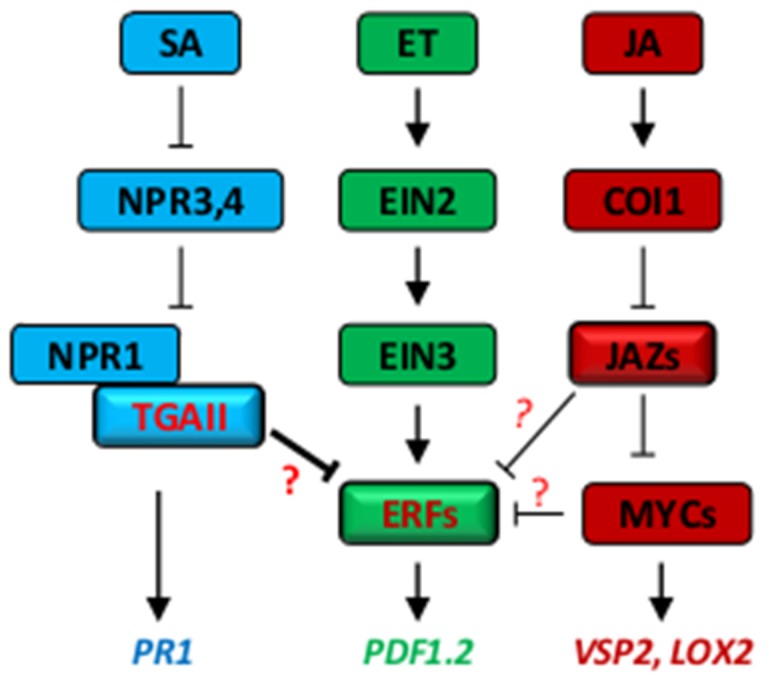
References
-
- Thomma B.P., Eggermont K., Penninckx I.A., Mauch-Mani B., Vogelsang R., Cammue B.P., Broekaert W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA. 1998;95:15107–15111. doi: 10.1073/pnas.95.25.15107. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
