

SPR-Measured Dissociation Kinetics of PROTAC Ternary Complexes Influence Target Degradation Rate
- PMID: 30721025
- PMCID: PMC6423499
- DOI: 10.1021/acschembio.9b00092
SPR-Measured Dissociation Kinetics of PROTAC Ternary Complexes Influence Target Degradation Rate
Abstract
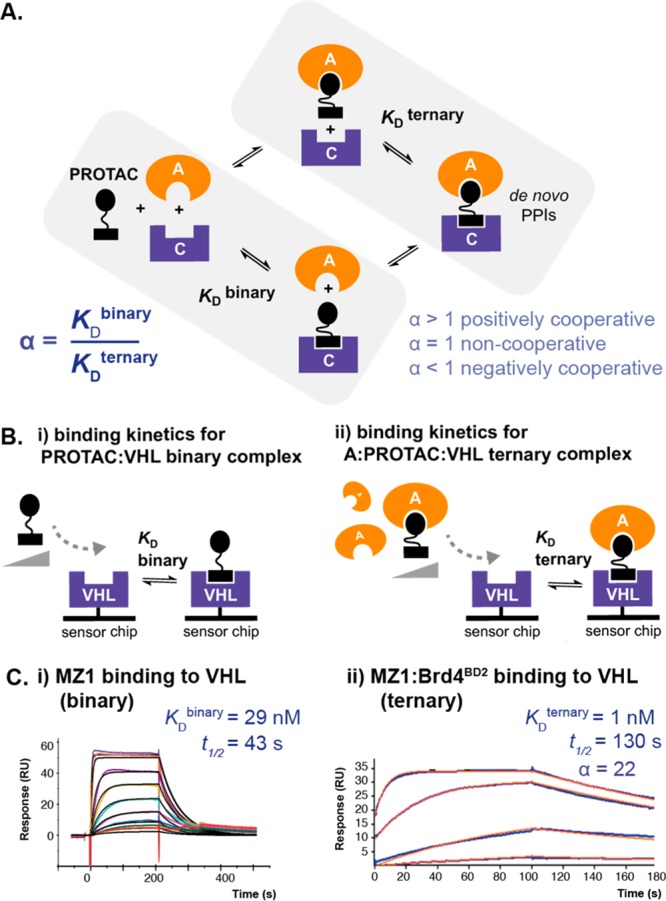
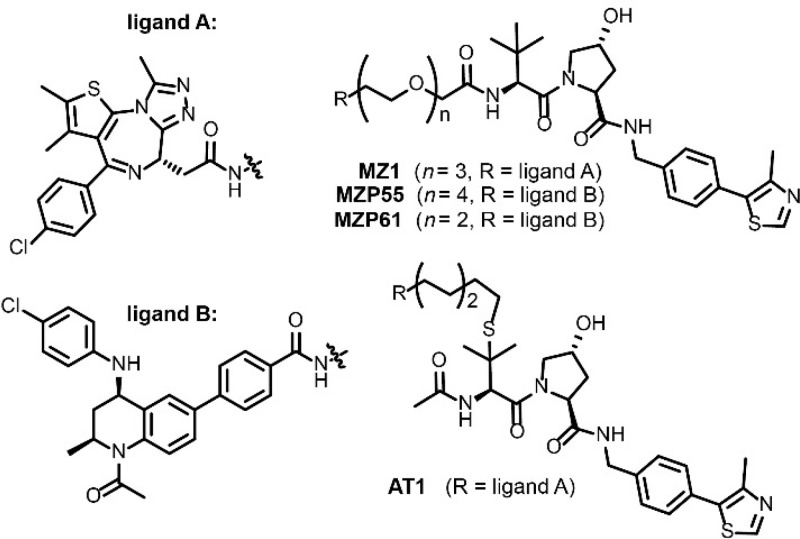
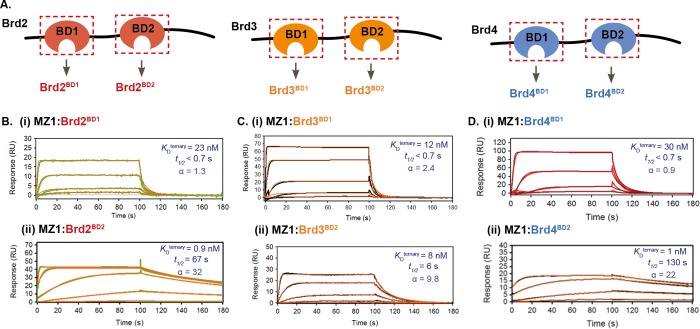
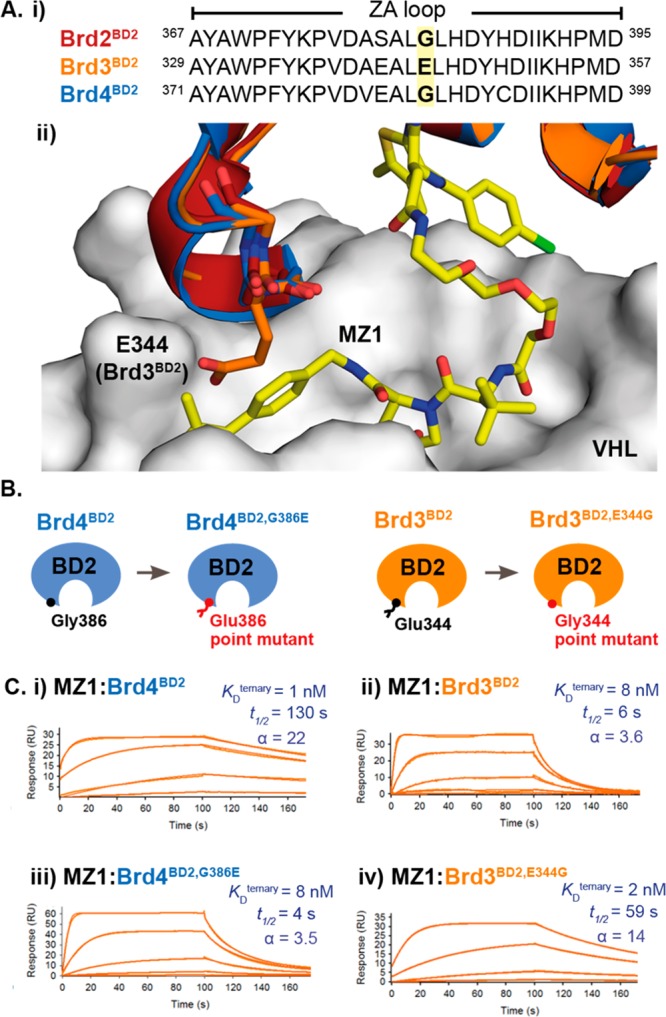
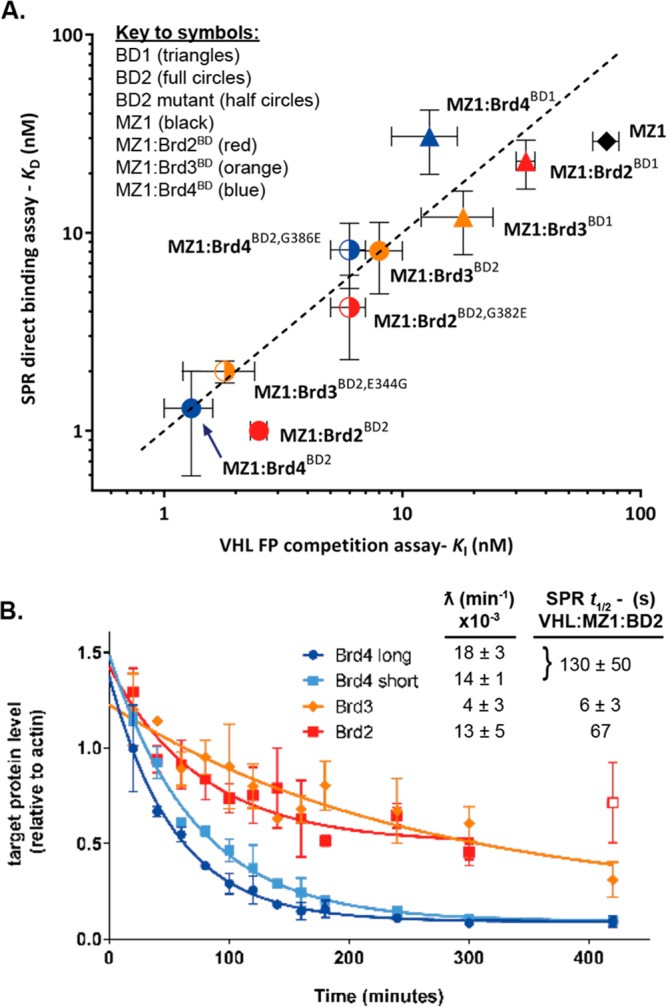
Bifunctional degrader molecules, known as proteolysis-targeting chimeras (PROTACs), function by recruiting a target to an E3 ligase, forming a target/PROTAC/ligase ternary complex. Despite the importance of this key intermediate species, no detailed validation of a method to directly determine binding parameters for ternary complex kinetics has been reported, and it remains to be addressed whether tuning the kinetics of PROTAC ternary complexes may be an effective strategy to improve the efficiency of targeted protein degradation. Here, we develop an SPR-based assay to quantify the stability of PROTAC-induced ternary complexes by measuring for the first time the kinetics of their formation and dissociation in vitro using purified proteins. We benchmark our assay using four PROTACs that target the bromodomains (BDs) of bromodomain and extraterminal domain proteins Brd2, Brd3, and Brd4 to the von Hippel-Lindau E3 ligase (VHL). We reveal marked differences in ternary complex off-rates for different PROTACs that exhibit either positive or negative cooperativity for ternary complex formation relative to binary binding. The positively cooperative degrader MZ1 forms comparatively stable and long-lived ternary complexes with either Brd4BD2 or Brd2BD2 and VHL. Equivalent complexes with Brd3BD2 are destabilized due to a single amino acid difference (Glu/Gly swap) present in the bromodomain. We observe that this difference in ternary complex dissociative half-life correlates to a greater initial rate of intracellular degradation of Brd2 and Brd4 relative to Brd3. These findings establish a novel assay to measure the kinetics of PROTAC ternary complexes and elucidate the important kinetic parameters that drive effective target degradation.
Conflict of interest statement
The authors declare the following competing financial interest(s): The Ciulli laboratory receives sponsored research support from Boehringer Ingelheim and Nurix, Inc. A.C. is a scientific founder, director, and shareholder of Amphista Therapeutics, a company that is developing targeted protein degradation therapeutic platforms.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
