

Prickle1 is required for EMT and migration of zebrafish cranial neural crest
- PMID: 30721665
- PMCID: PMC6480417
- DOI: 10.1016/j.ydbio.2019.01.018
Prickle1 is required for EMT and migration of zebrafish cranial neural crest
Abstract
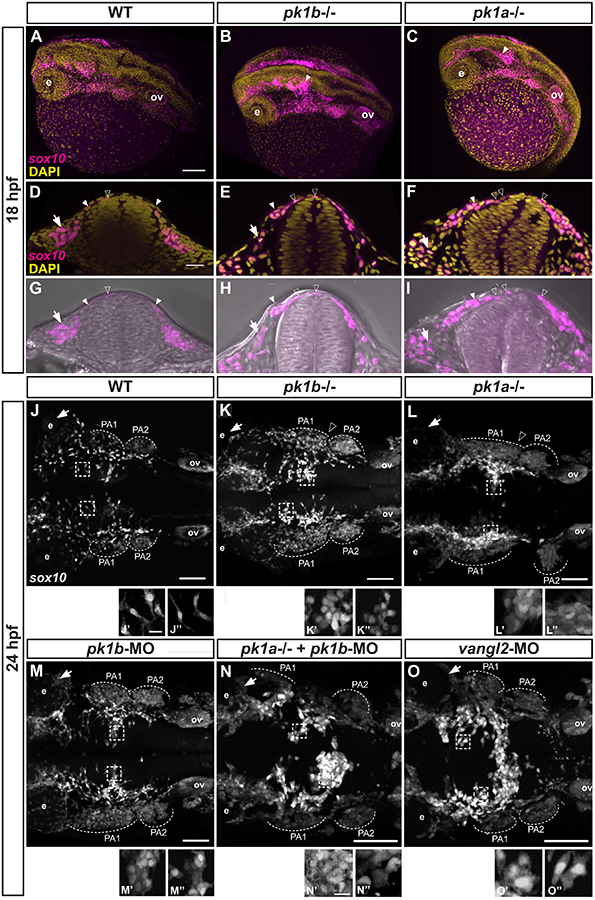
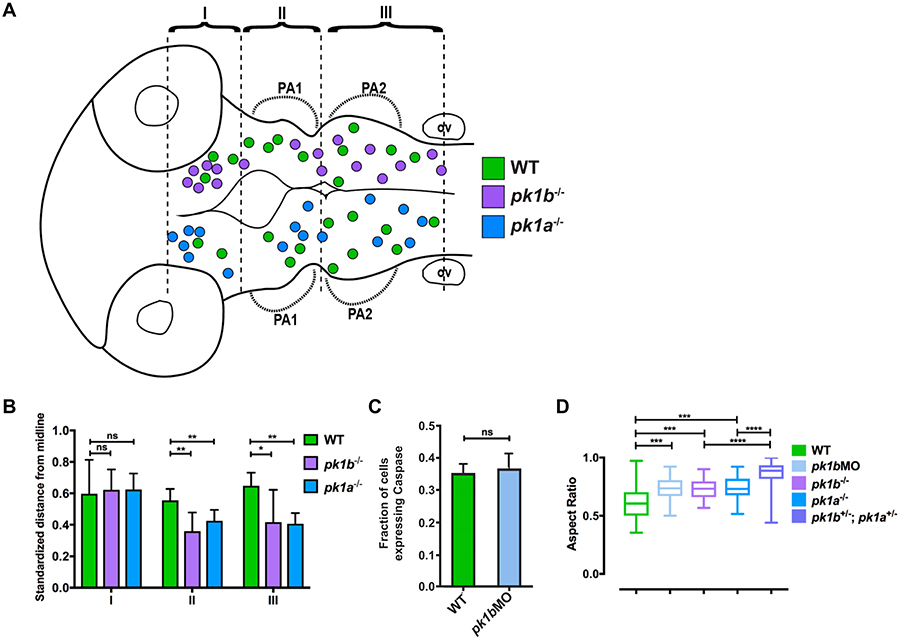
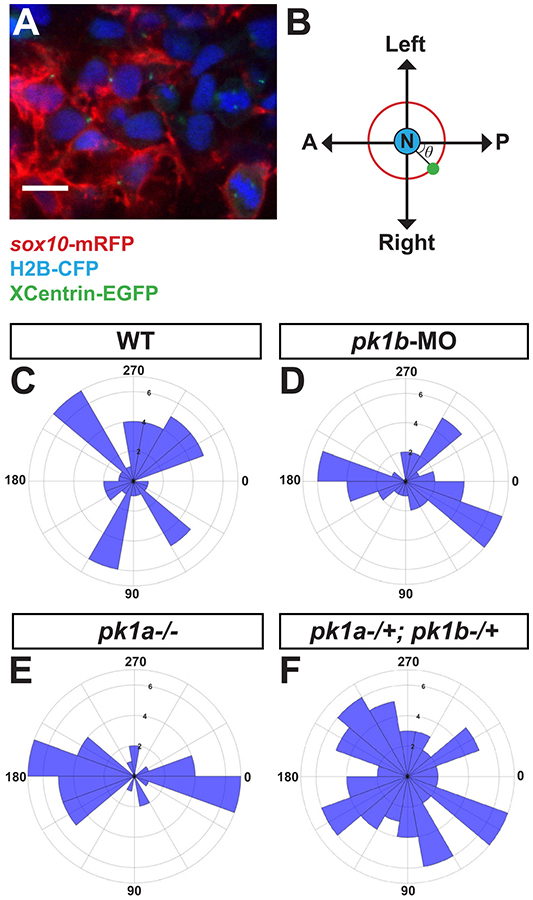
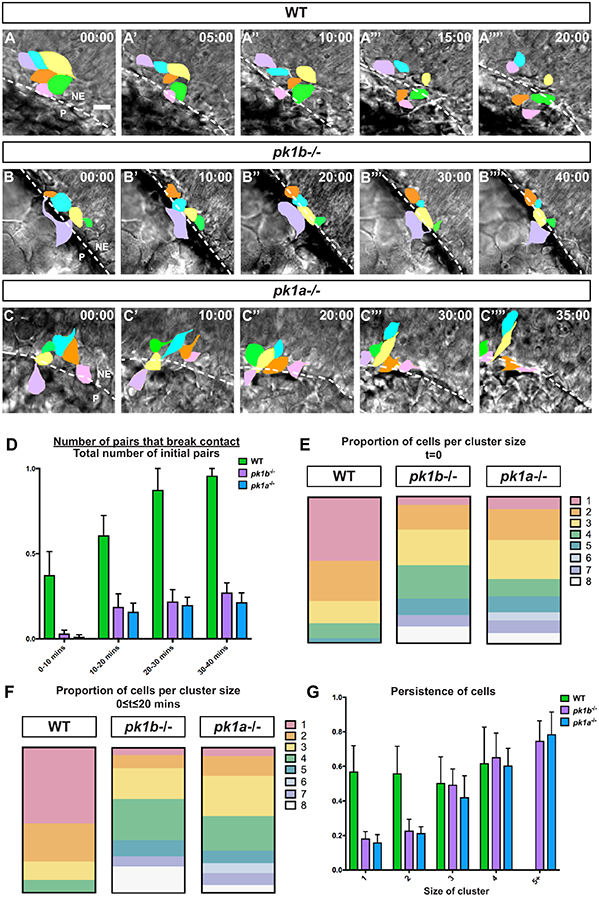
The neural crest-a key innovation of the vertebrates-gives rise to diverse cell types including melanocytes, neurons and glia of the peripheral nervous system, and chondrocytes of the jaw and skull. Proper development of the cephalic region is dependent on the tightly-regulated specification and migration of cranial neural crest cells (NCCs). The core PCP proteins Frizzled and Disheveled have previously been implicated in NCC migration. Here we investigate the functions of the core PCP proteins Prickle1a and Prickle1b in zebrafish cranial NCC development. Using analysis of pk1a and pk1b mutant embryos, we uncover similar roles for both genes in facilitating cranial NCC migration. Disruption of either gene causes pre-migratory NCCs to cluster together at the dorsal aspect of the neural tube, where they adopt aberrant polarity and movement. Critically, in investigating Pk1-deficient cells that fail to migrate ventrolaterally, we have also uncovered roles for pk1a and pk1b in the epithelial-to-mesenchymal transition (EMT) of pre-migratory NCCs that precedes their collective migration to the periphery. Normally, during EMT, pre-migratory NCCs transition from a neuroepithelial to a bleb-based and subsequently, mesenchymal morphology capable of directed migration. When either Pk1a or Pk1b is disrupted, NCCs continue to perform blebbing behaviors characteristic of pre-migratory cells over extended time periods, indicating a block in a key transition during EMT. Although some Pk1-deficient NCCs transition successfully to mesenchymal, migratory morphologies, they fail to separate from neighboring NCCs. Additionally, Pk1b-deficient NCCs show elevated levels of E-Cadherin and reduced levels of N-Cadherin, suggesting that Prickle1 molecules regulate Cadherin levels to ensure the completion of EMT and the commencement of cranial NCC migration. We conclude that Pk1 plays crucial roles in cranial NCCs both during EMT and migration. These roles are dependent on the regulation of E-Cad and N-Cad.
Keywords: Cadherin; Cranial neural crest; EMT; Planar cell polarity; Prickle; Zebrafish.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures





References
-
- Andrieu C, Montigny A, Alfandari D, and Theveneau E (2018). Basolateral localization of MMP14 drives apicobasal polarity change during EMT independently of its catalytic activity. bioRxiv, 402180.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
