

A Disintegrin and Metalloproteinase 9 Domain (ADAM9) Is a Major Susceptibility Factor in the Early Stages of Encephalomyocarditis Virus Infection
- PMID: 30723129
- PMCID: PMC6428755
- DOI: 10.1128/mBio.02734-18
A Disintegrin and Metalloproteinase 9 Domain (ADAM9) Is a Major Susceptibility Factor in the Early Stages of Encephalomyocarditis Virus Infection
Abstract
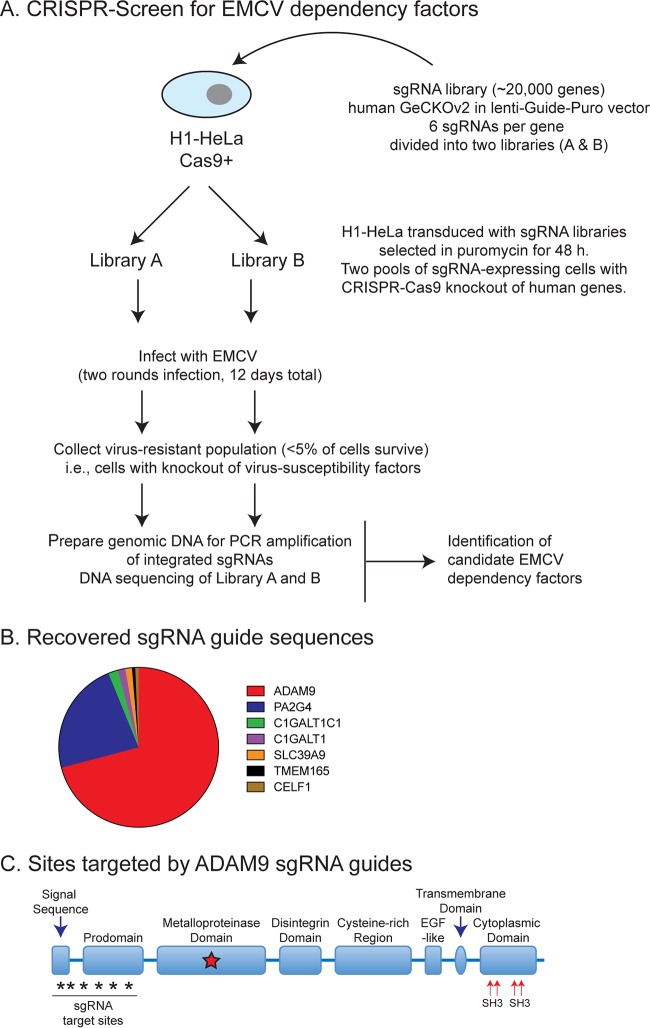
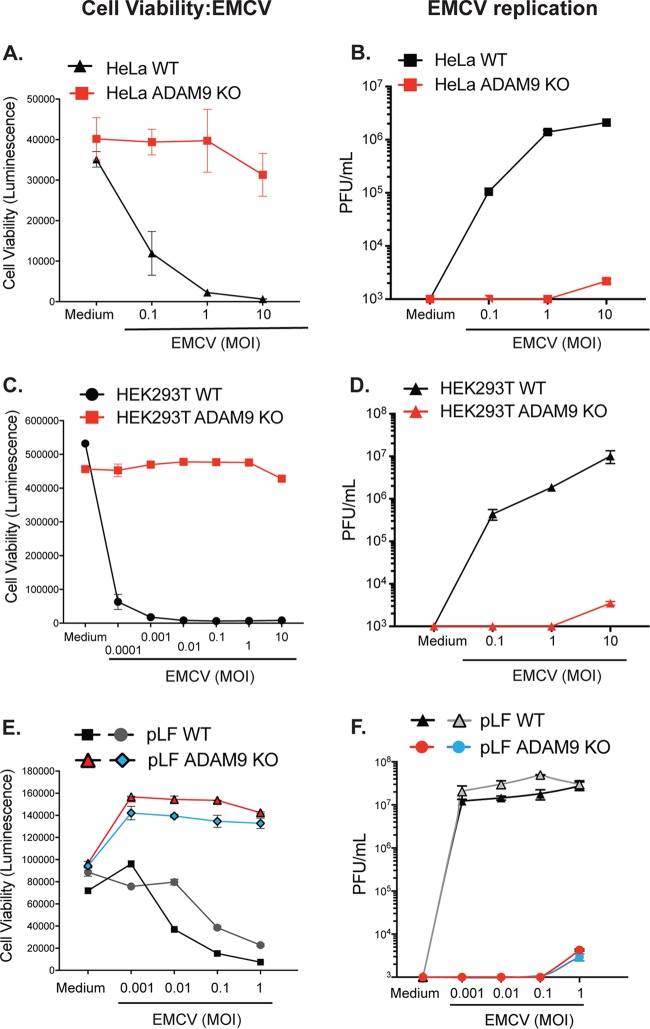
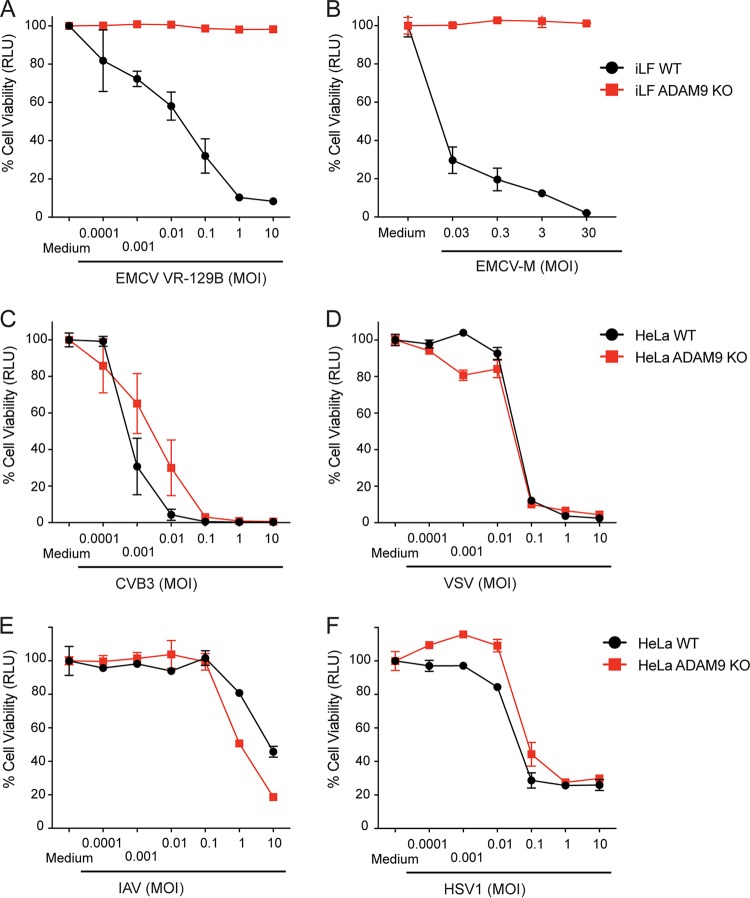
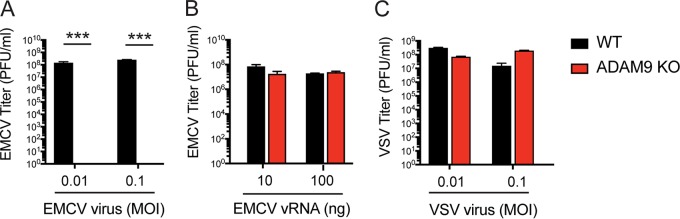
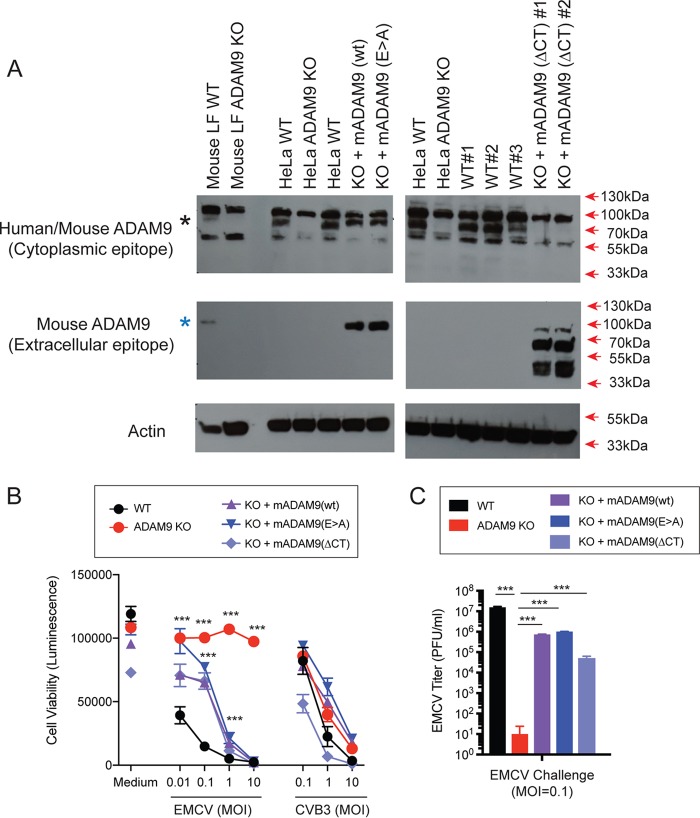
Encephalomyocarditis virus (EMCV) is a picornavirus that produces lytic infections in murine and human cells. Employing a genome-wide CRISPR-Cas9 knockout screen to find host factors required for EMCV infection, we identified a role for ADAM9 in EMCV infection. CRISPR-mediated deletion of ADAM9 in multiple human cell lines rendered the cells highly resistant to EMCV infection and cell death. Primary fibroblasts from ADAM9 KO mice were also strongly resistant to EMCV infection and cell death. In contrast, ADAM9 KO and WT cells were equally susceptible to infection with other viruses, including the picornavirus Coxsackie virus B. ADAM9 KO cells failed to produce viral progeny when incubated with EMCV. However, bypassing EMCV entry into cells through delivery of viral RNA directly to the cytosol yielded infectious EMCV virions from ADAM9 KO cells, suggesting that ADAM9 is not required for EMCV replication post-entry. These findings establish that ADAM9 is required for the early stage of EMCV infection, likely for virus entry or viral genome delivery to the cytosol.IMPORTANCE Viral myocarditis is a leading cause of death in the United States, contributing to numerous unexplained deaths in people ≤35 years old. Enteroviruses contribute to many cases of human myocarditis. Encephalomyocarditis virus (EMCV) infection causes viral myocarditis in rodent models, but its receptor requirements have not been fully identified. CRISPR-Cas9 screens can identify host dependency factors essential for EMCV infection and enhance our understanding of key events that follow viral infection, potentially leading to new strategies for preventing viral myocarditis. Using a CRISPR-Cas9 screen, we identified
Keywords: a disintegrin and metalloproteinase 9 domain (ADAM9); encephalomyocarditis virus; functional genomic screen.
Copyright © 2019 Bazzone et al.
Figures
Comment in
-
Encephalomyocarditis Virus Entry Unveiled.mBio. 2019 Mar 26;10(2):e00305-19. doi: 10.1128/mBio.00305-19. mBio. 2019. PMID: 30914507 Free PMC article.
References
-
- Murnane TG, Craighead JE, Mondragon H, Shelokov A. 1960. Fatal disease of swine due to encephalomyocarditis virus. Science 131:498–499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
