

A Genome-wide Haploid Genetic Screen Identifies Regulators of Glutathione Abundance and Ferroptosis Sensitivity
- PMID: 30726737
- PMCID: PMC6424331
- DOI: 10.1016/j.celrep.2019.01.043
A Genome-wide Haploid Genetic Screen Identifies Regulators of Glutathione Abundance and Ferroptosis Sensitivity
Abstract
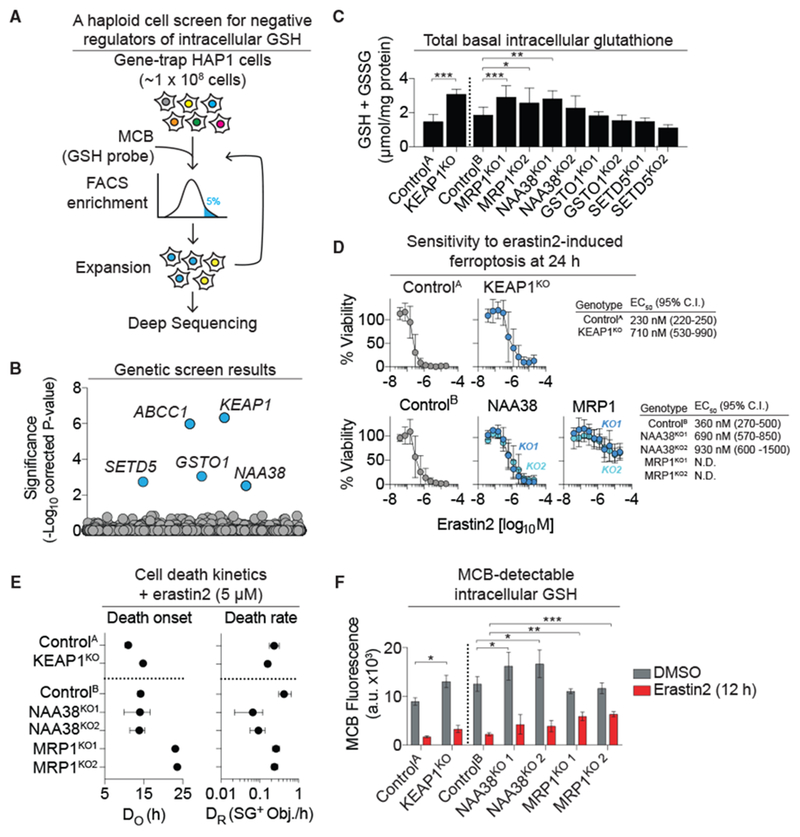
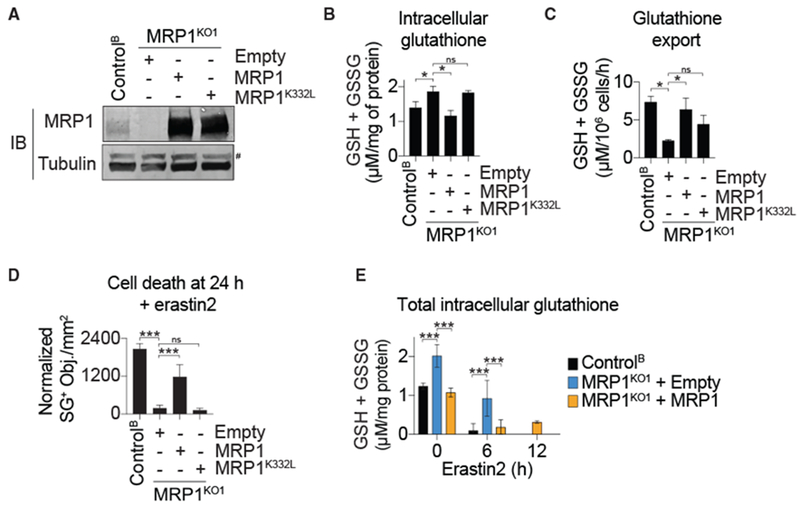
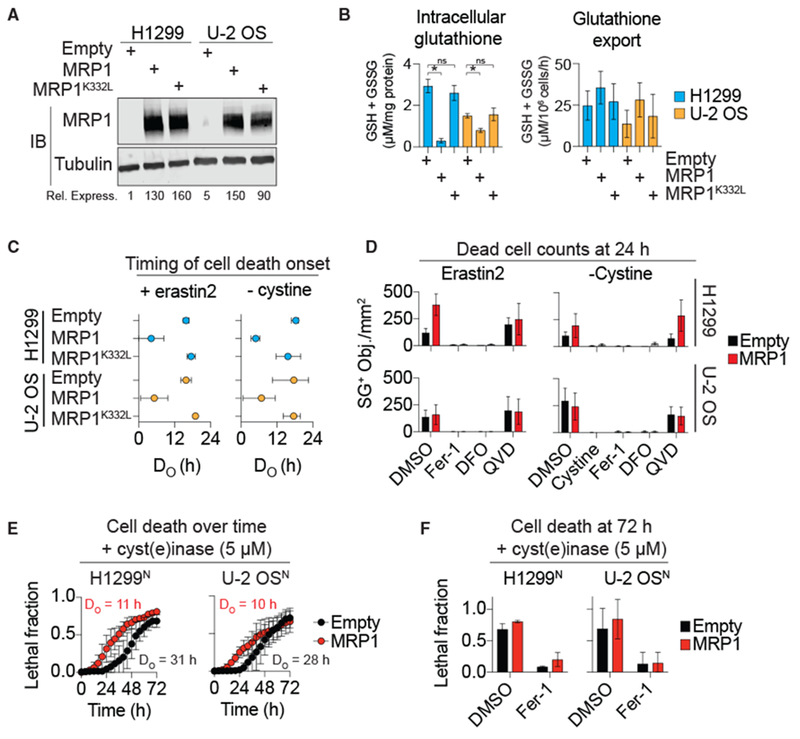
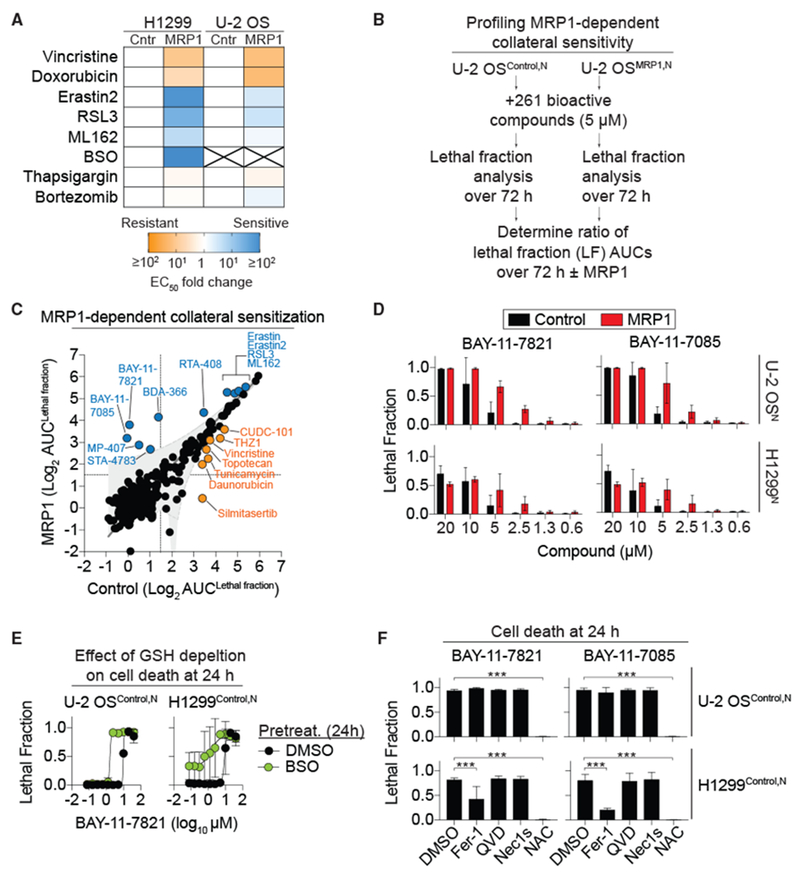
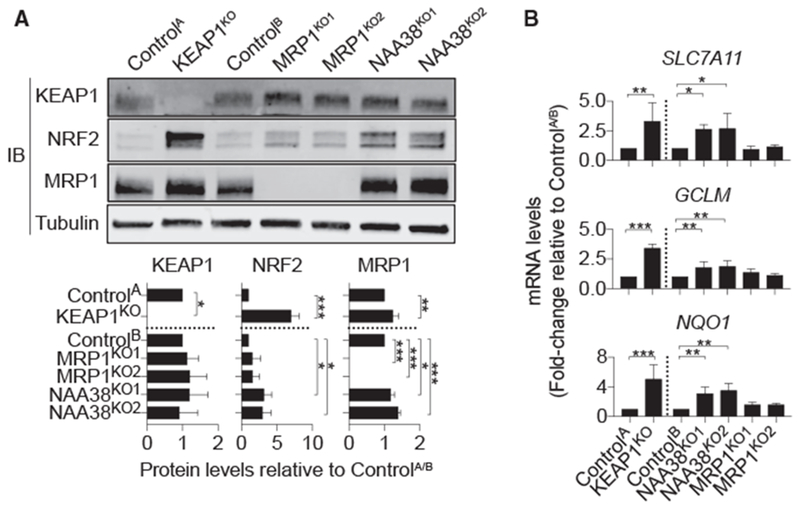
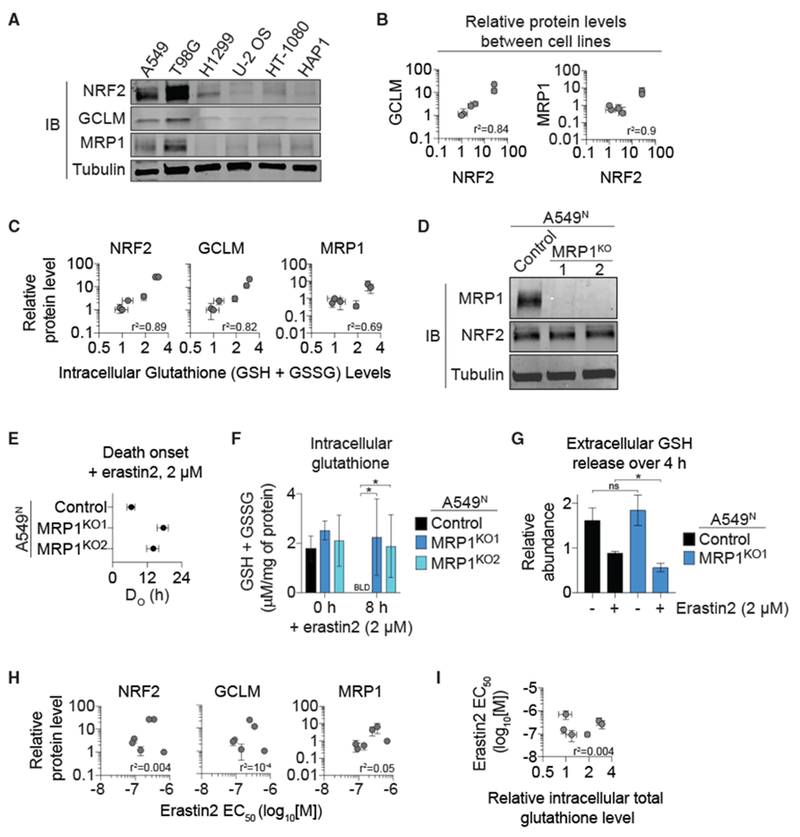
The tripeptide glutathione suppresses the iron-dependent, non-apoptotic cell death process of ferroptosis. How glutathione abundance is regulated in the cell and how this regulation alters ferroptosis sensitivity is poorly understood. Using genome-wide human haploid genetic screening technology coupled to fluorescence-activated cell sorting (FACS), we directly identify genes that regulate intracellular glutathione abundance and characterize their role in ferroptosis regulation. Disruption of the ATP binding cassette (ABC)-family transporter multidrug resistance protein 1 (MRP1) prevents glutathione efflux from the cell and strongly inhibits ferroptosis. High levels of MRP1 expression decrease sensitivity to certain pro-apoptotic chemotherapeutic drugs, while collaterally sensitizing to all tested pro-ferroptotic agents. By contrast, disruption of KEAP1 and NAA38, leading to the stabilization of the transcription factor NRF2, increases glutathione levels but only weakly protects from ferroptosis. This is due in part to concomitant NRF2-mediated upregulation of MRP1. These results pinpoint glutathione efflux as an unanticipated regulator of ferroptosis sensitivity.
Keywords: ROS; cancer; collateral sensitivity; ferroptosis; glutathione; iron; metabolite efflux; multidrug resistance; necrosis.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Aksnes H, Hole K, and Arnesen T (2015). Molecular, cellular, and physiological significance of N-terminal acetylation. Int. Rev. Cell Mol. Biol 316, 267–305. - PubMed
-
- Chang L-C, Chiang S-K, Chen S-E, Yu Y-L, Chou R-H, and Chang W-C (2018). Heme oxygenase-1 mediates BAY 11-7085 induced ferroptosis. Cancer Lett. 416, 124–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
