

Differential Antibody-Based Immune Response against Isolated GP1 Receptor-Binding Domains from Lassa and Junín Viruses
- PMID: 30728269
- PMCID: PMC6450128
- DOI: 10.1128/JVI.00090-19
Differential Antibody-Based Immune Response against Isolated GP1 Receptor-Binding Domains from Lassa and Junín Viruses
Abstract
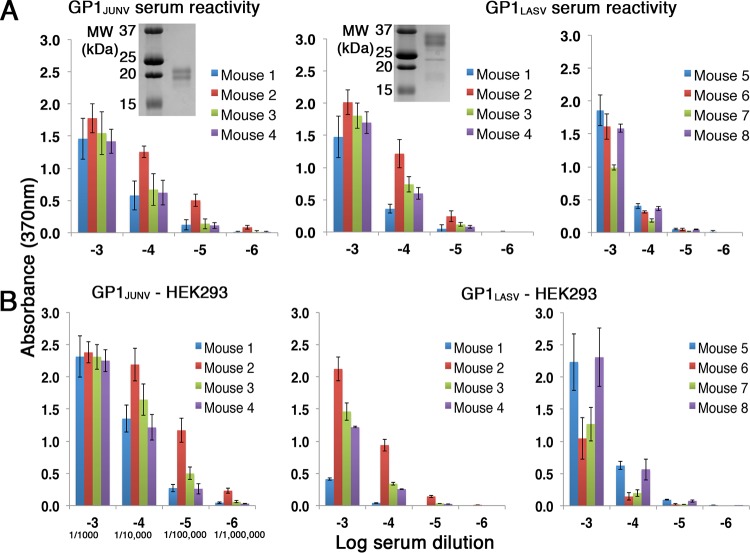
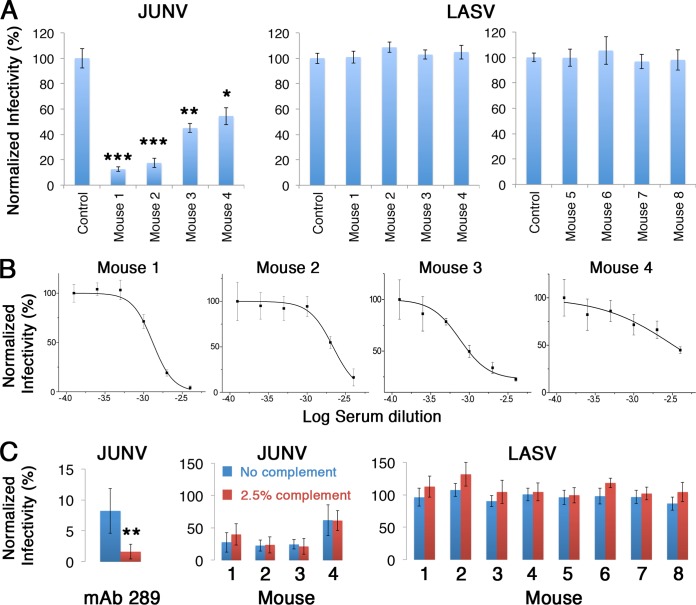
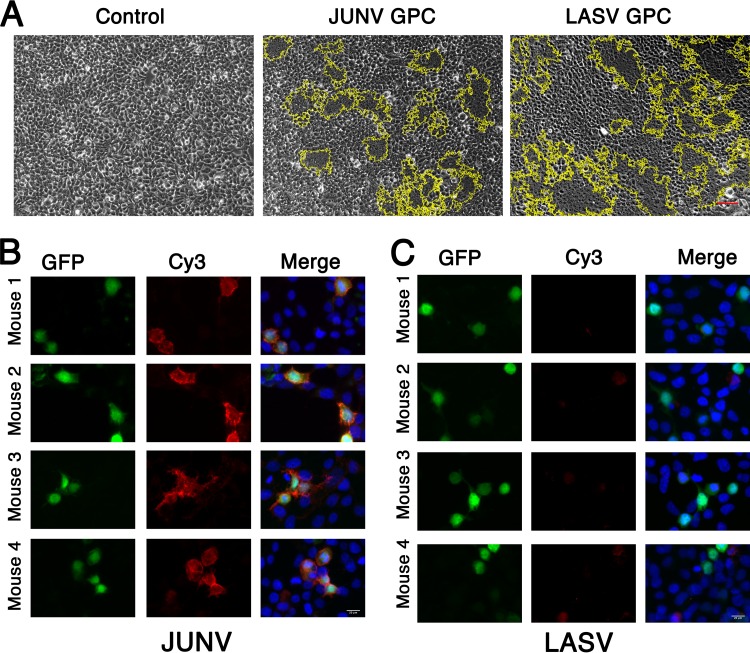
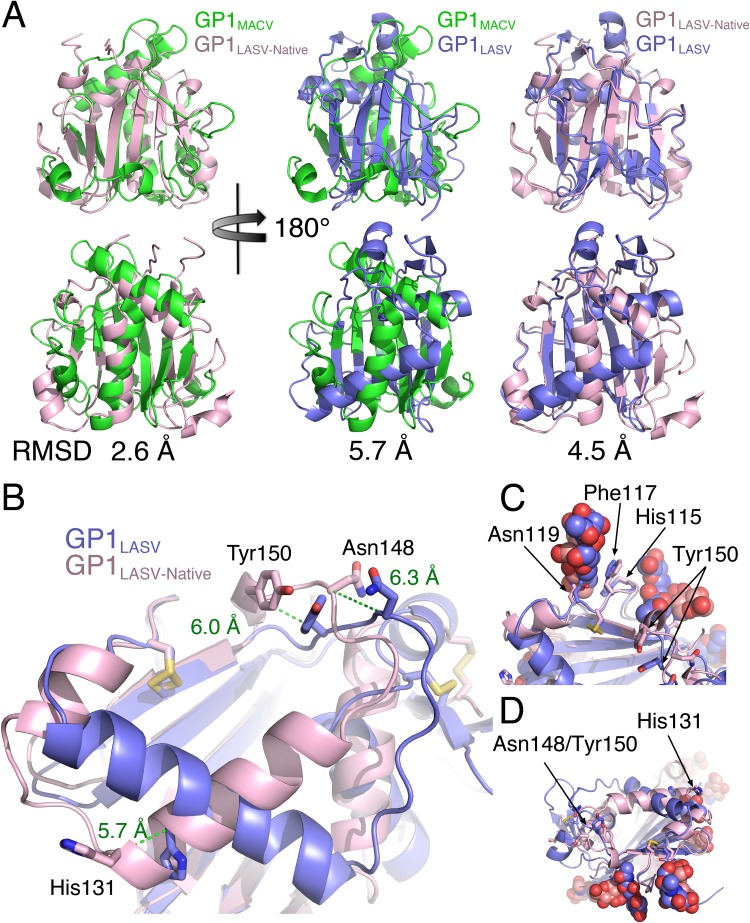
There are two predominant subgroups in the Arenaviridae family of viruses, the Old World and the New World viruses, that use distinct cellular receptors for entry. While New World viruses typically elicit good neutralizing antibody responses, the Old World viruses generally evade such responses. Antibody-based immune responses are directed against the glycoprotein spike complexes that decorate the viruses. A thick coat of glycans reduces the accessibility of antibodies to the surface of spike complexes from Old World viruses, but other mechanisms may further hamper the development of efficient humoral responses. Specifically, it was suggested that the GP1 receptor-binding module of the Old World Lassa virus might help with evasion of the humoral response. Here we investigated the immunogenicity of the GP1 domain from Lassa virus and compared it to that of the GP1 domain from the New World Junín virus. We found striking differences in the ability of antibodies that were developed against these immunogens to target the same GP1 receptor-binding domains in the context of the native spike complexes. Whereas GP1 from Junín virus elicited productive neutralizing responses, GP1 from Lassa virus elicited only nonproductive responses. These differences can be rationalized by the conformational changes that GP1 from Lassa virus but not GP1 from Junín virus undergoes after dissociating from the trimeric spike complex. Hence, shedding of GP1 in the case of Lassa virus can indeed serve as a mechanism to subvert the humoral immune response. Moreover, the realization that a recombinant protein may be used to elicit a productive response against the New World Junín virus may suggest a novel and safe way to design future vaccines.IMPORTANCE Some viruses that belong to the Arenaviridae family, like Lassa and Junín viruses, are notorious human pathogens, which may lead to fatal outcomes when they infect people. It is thus important to develop means to combat these viruses. For developing effective vaccines, it is vital to understand the basic mechanisms that these viruses utilize in order to evade or overcome host immune responses. It was previously noted that the GP1 receptor-binding domain from Lassa virus is shed and accumulates in the serum of infected individuals. This raised the possibility that Lassa virus GP1 may function as an immunological decoy. Here we demonstrate that mice develop nonproductive immune responses against GP1 from Lassa virus, which is in contrast to the effective neutralizing responses that GP1 from Junín virus elicits. Thus, GP1 from Lassa virus is indeed an immunological decoy and GP1 from Junín virus may serve as a constituent of a future vaccine.
Keywords: arenavirus; immune response; vaccines.
Copyright © 2019 American Society for Microbiology.
Figures
References
-
- Oldstone MB. 2002. Arenaviruses. II. The molecular pathogenesis of arenavirus infections. Introduction. Curr Top Microbiol Immunol 263:V–XII. - PubMed
-
- Oldstone MB. 2002. Arenaviruses. I. The epidemiology molecular and cell biology of arenaviruses. Introduction. Curr Top Microbiol Immunol 262:V–XII. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
