

Z-DNA and Z-RNA in human disease
- PMID: 30729177
- PMCID: PMC6323056
- DOI: 10.1038/s42003-018-0237-x
Z-DNA and Z-RNA in human disease
Abstract
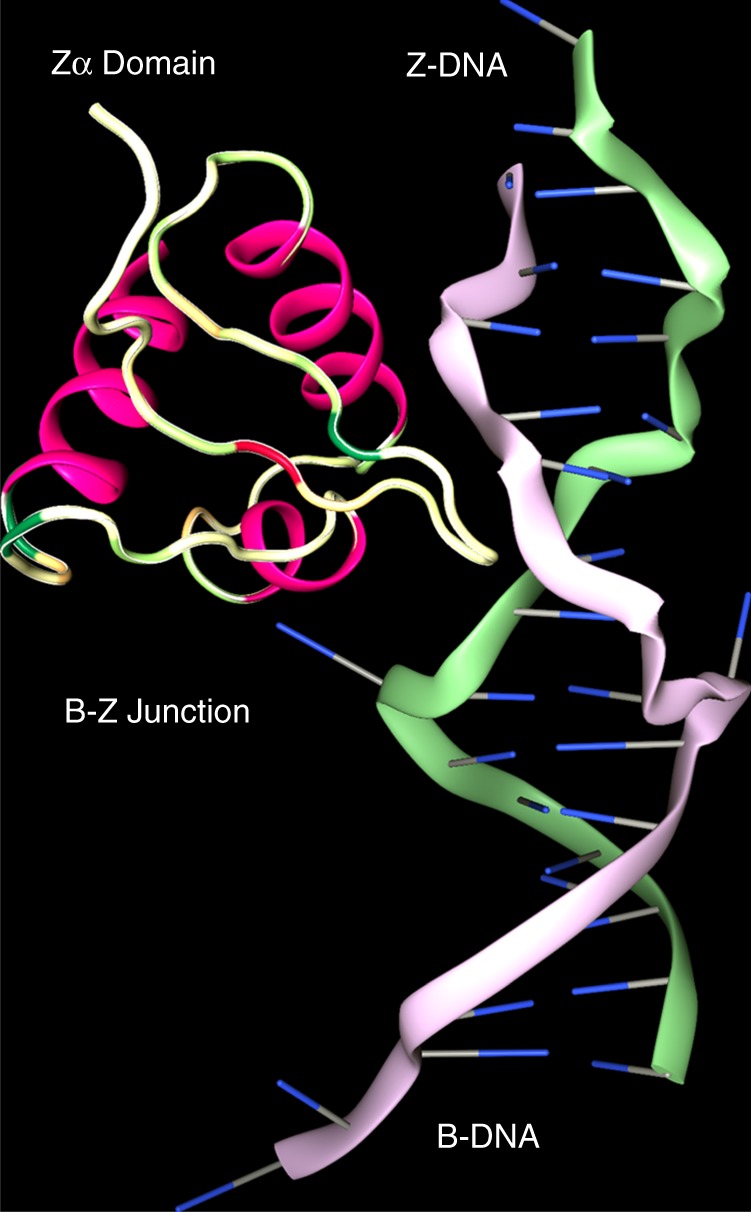
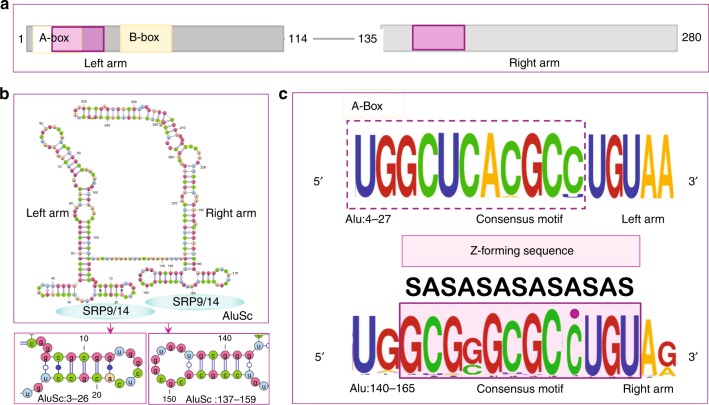
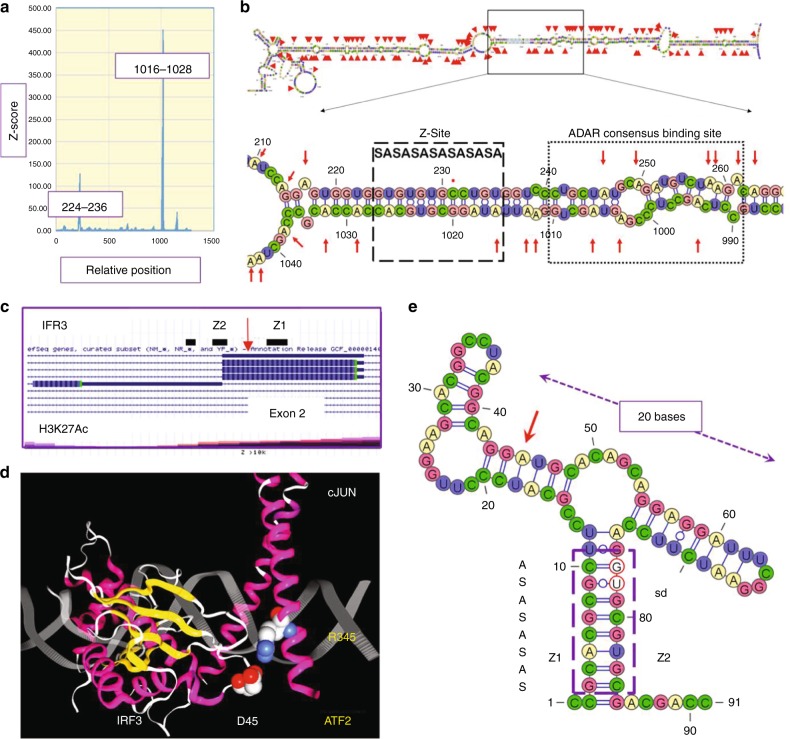
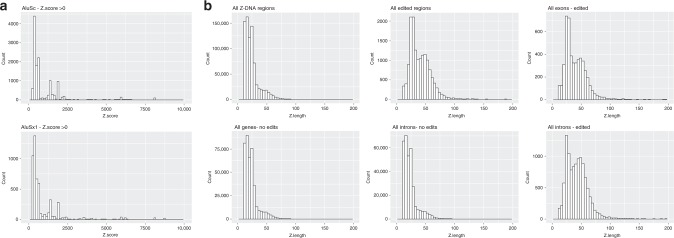
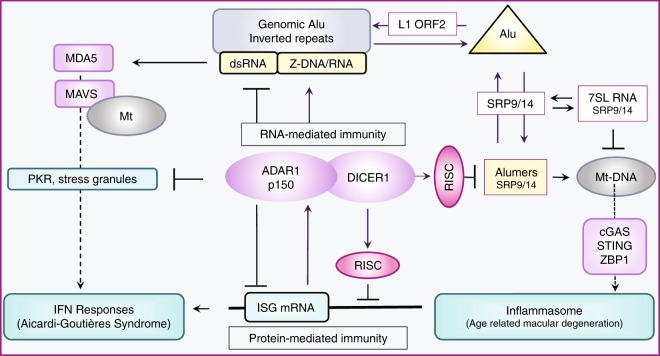
Left-handed Z-DNA/Z-RNA is bound with high affinity by the Zα domain protein family that includes ADAR (a double-stranded RNA editing enzyme), ZBP1 and viral orthologs regulating innate immunity. Loss-of-function mutations in ADAR p150 allow persistent activation of the interferon system by Alu dsRNAs and are causal for Aicardi-Goutières Syndrome. Heterodimers of ADAR and DICER1 regulate the switch from RNA- to protein-centric immunity. Loss of DICER1 function produces age-related macular degeneration, a different type of Alu-mediated disease. The overlap of Z-forming sites with those for the signal recognition particle likely limits invasion of primate genomes by Alu retrotransposons.
Conflict of interest statement
The author declares no competing interests.
Figures
References
-
- Patterson JB, Thomis DC, Hans SL, Samuel CE. Mechanism of interferon action: double-stranded RNA-specific adenosine deaminase from human cells is inducible by alpha and gamma interferons. Virology. 1995;210:508–511. - PubMed
-
- Schade M, et al. A 6 bp Z-DNA hairpin binds two Z alpha domains from the human RNA editing enzyme ADAR1. FEBS Lett. 1999;458:27–31. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
