

Mitotic chromosome alignment ensures mitotic fidelity by promoting interchromosomal compaction during anaphase
- PMID: 30733233
- PMCID: PMC6446859
- DOI: 10.1083/jcb.201807228
Mitotic chromosome alignment ensures mitotic fidelity by promoting interchromosomal compaction during anaphase
Abstract
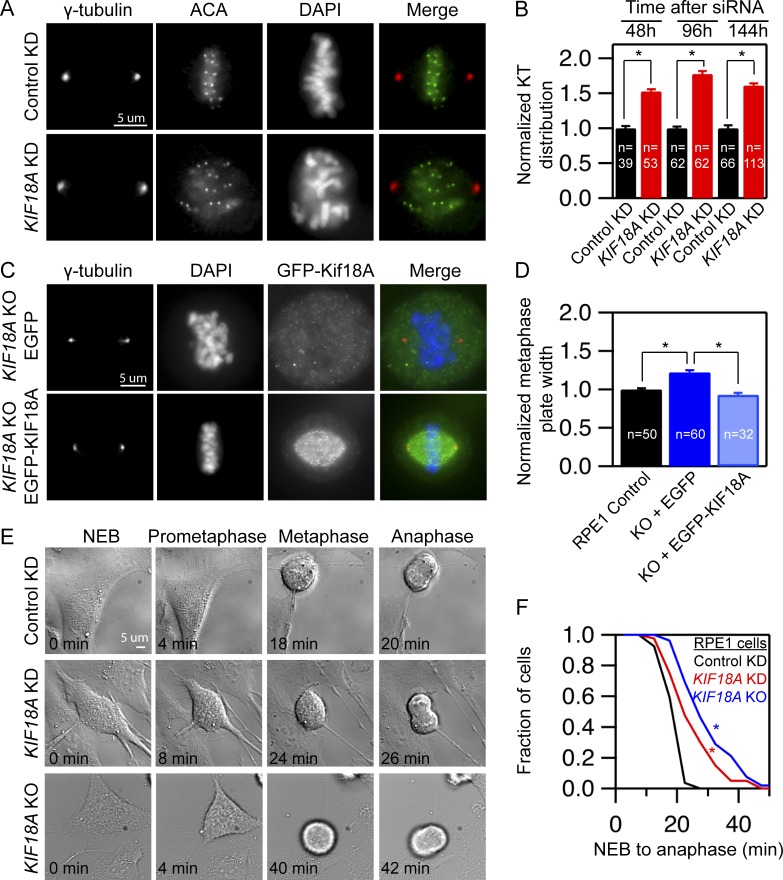
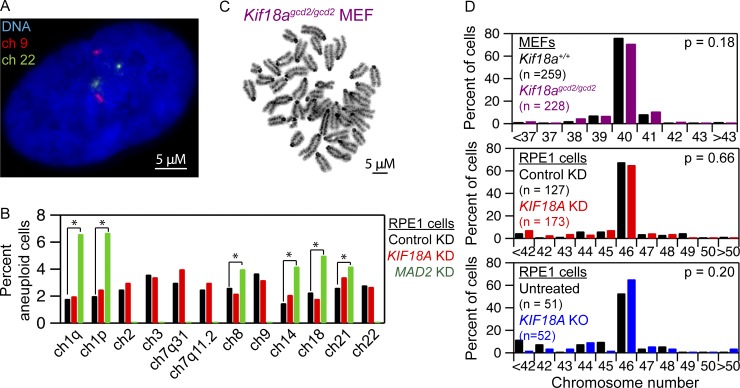
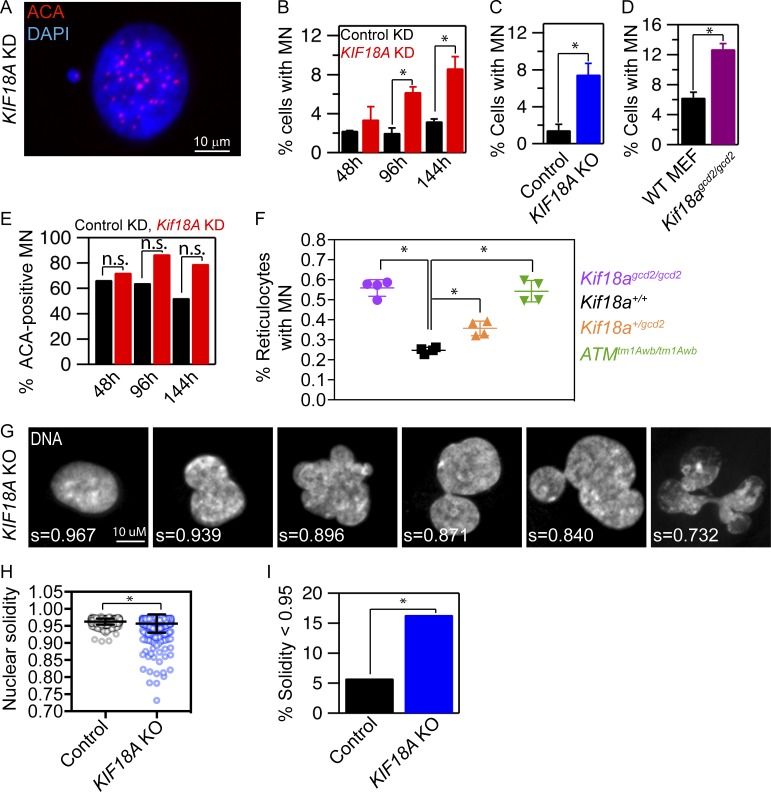
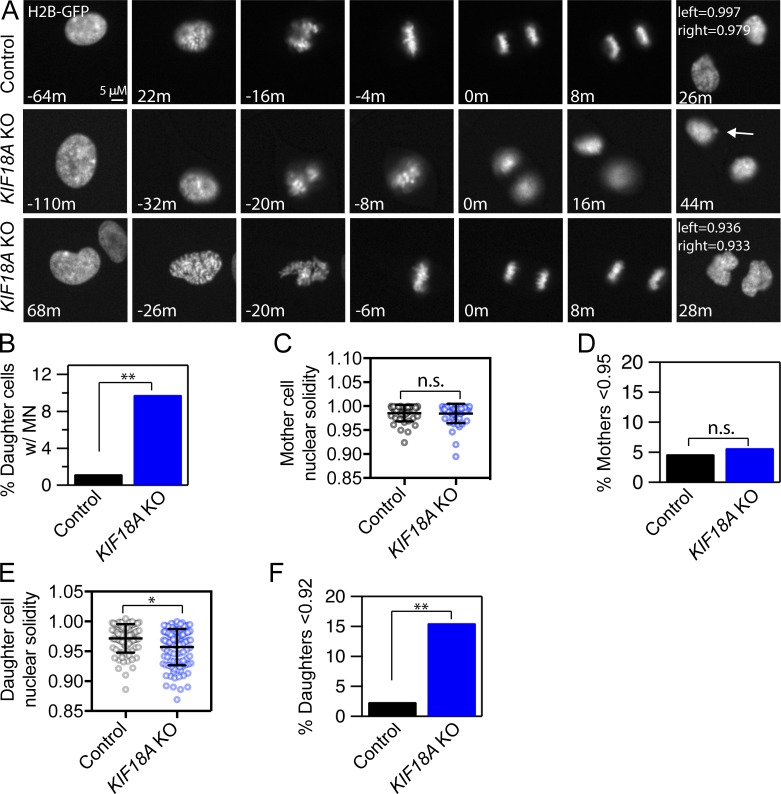
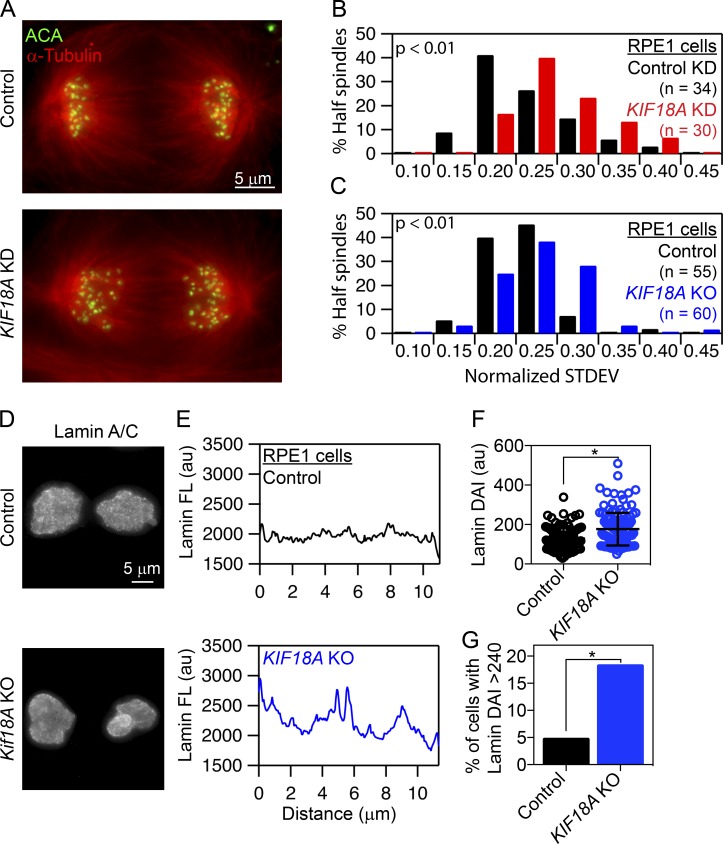
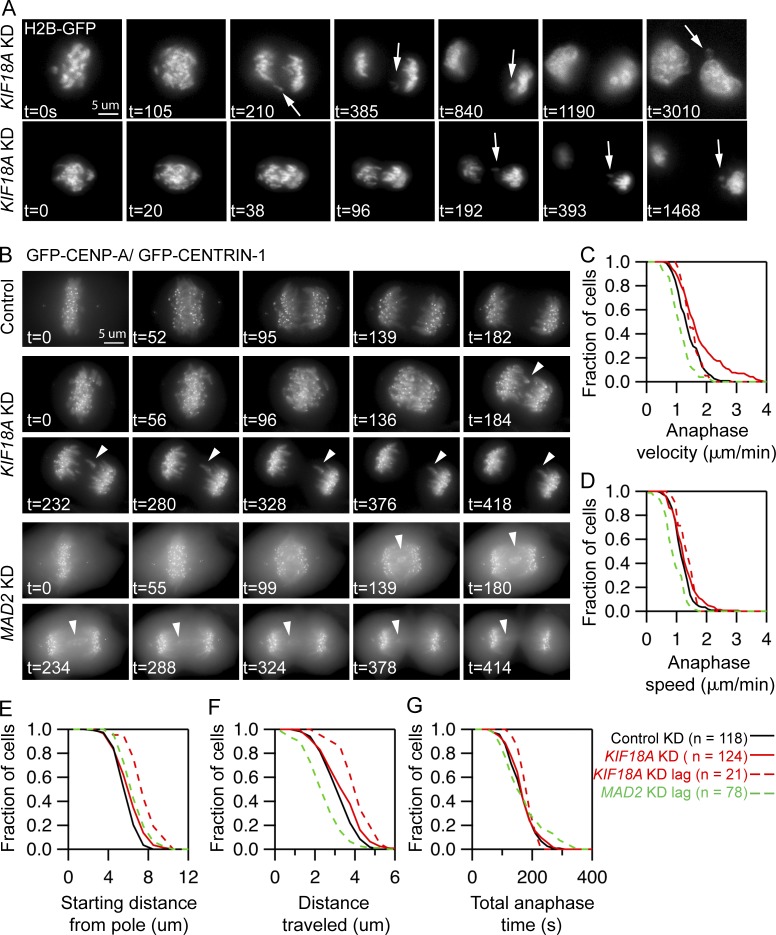
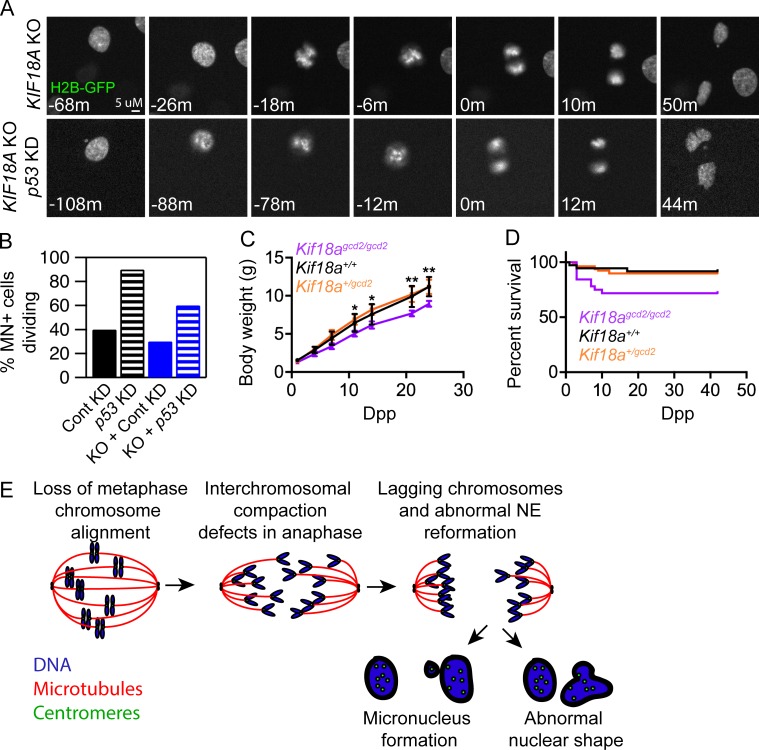
Chromosome alignment at the equator of the mitotic spindle is a highly conserved step during cell division; however, its importance to genomic stability and cellular fitness is not understood. Normal mammalian somatic cells lacking KIF18A function complete cell division without aligning chromosomes. These alignment-deficient cells display normal chromosome copy numbers in vitro and in vivo, suggesting that chromosome alignment is largely dispensable for maintenance of euploidy. However, we find that loss of chromosome alignment leads to interchromosomal compaction defects during anaphase, abnormal organization of chromosomes into a single nucleus at mitotic exit, and the formation of micronuclei in vitro and in vivo. These defects slow cell proliferation and are associated with impaired postnatal growth and survival in mice. Our studies support a model in which the alignment of mitotic chromosomes promotes proper organization of chromosomes into a single nucleus and continued proliferation by ensuring that chromosomes segregate as a compact mass during anaphase.
© 2019 Fonseca et al.
Figures
Comment in
-
No chromosome left behind: The importance of metaphase alignment for mitotic fidelity.J Cell Biol. 2019 Apr 1;218(4):1086-1088. doi: 10.1083/jcb.201902041. Epub 2019 Mar 11. J Cell Biol. 2019. PMID: 30858193 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
