

Lnc-ing Trained Immunity to Chromatin Architecture
- PMID: 30733945
- PMCID: PMC6353842
- DOI: 10.3389/fcell.2019.00002
Lnc-ing Trained Immunity to Chromatin Architecture
Abstract
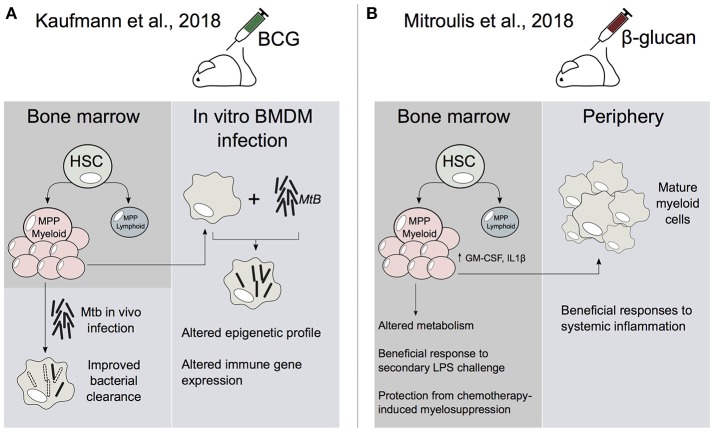
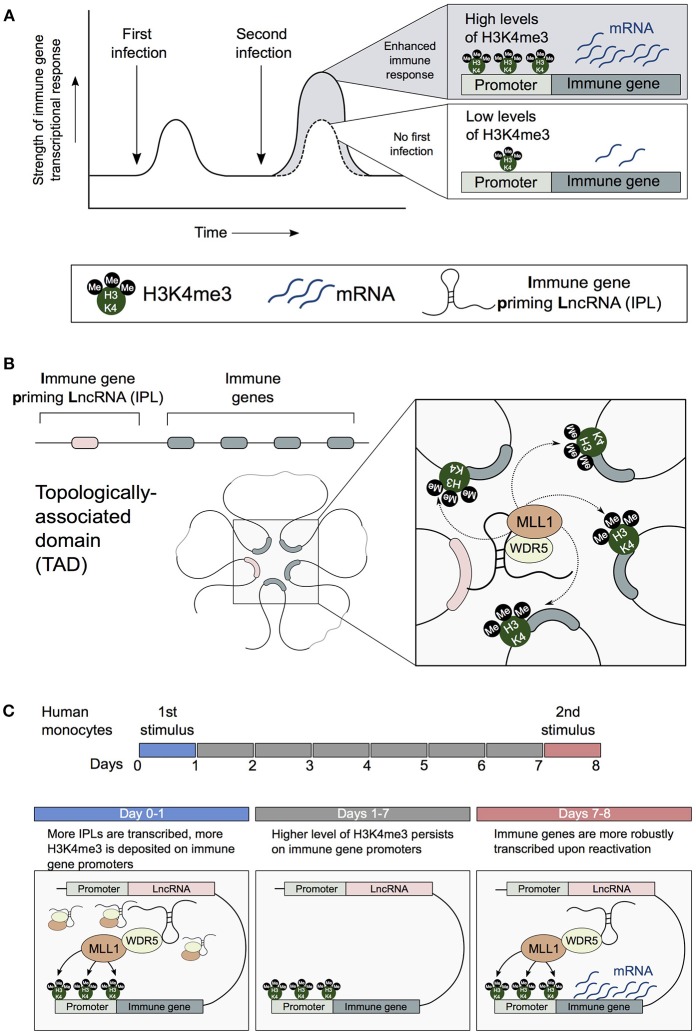
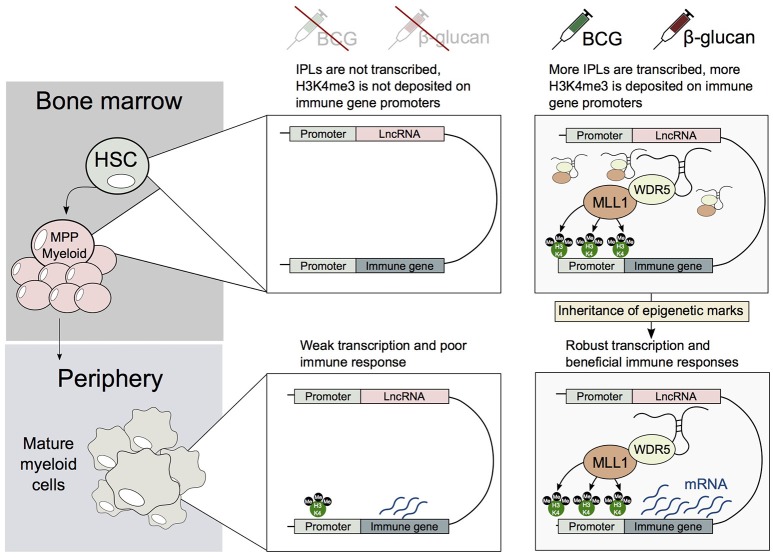
Human innate immune cells exposed to certain infections or stimuli develop enhanced immune responses upon re-infection with a different second stimulus, a process termed trained immunity. Recent studies have revealed that hematopoietic stem cells (HSCs) are integral to trained immune responses as they are able to "remember" transcriptional responses and transmit this state to their progeny to educate them how to respond to future infections. The macrophages that arise from trained HSCs are epigenetically reprogrammed and as a result robustly express immune genes, enhancing their capability to resolve infection. Accumulation of H3K4me3 epigenetic marks on multiple immune gene promoters underlie robust transcriptional responses during trained immune responses. However, the mechanism underpinning how these epigenetic marks accumulate at discrete immune gene loci has been poorly understood. In this review, we discuss the previously unexplored contributions of nuclear architecture and long non-coding RNAs on H3K4me3 promoter priming in trained immunity. Altering the activity of these lncRNAs presents a promising therapeutic approach to achieve immunomodulation in inflammatory disease states.
Keywords: BCG—Bacille Calmette-Guérin vaccine; LncRNA—long non-coding RNA; chromatin; epigenetic memory; innate immunity; macrophages; trained immunity; transcriptional (regulation).
Figures
References
-
- Cabezas-Wallscheid N., Klimmeck D., Hansson J., Lipka D. B., Reyes A., Wang Q., et al. . (2014). Identification of regulatory networks in HSCs and their immediate progeny via integrated proteome, transcriptome, and DNA methylome analysis. Cell Stem Cell 15, 507–522. 10.1016/j.stem.2014.07.005 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources