

Tight junction proteins at the blood-brain barrier: far more than claudin-5
- PMID: 30734065
- PMCID: PMC11105330
- DOI: 10.1007/s00018-019-03030-7
Tight junction proteins at the blood-brain barrier: far more than claudin-5
Abstract
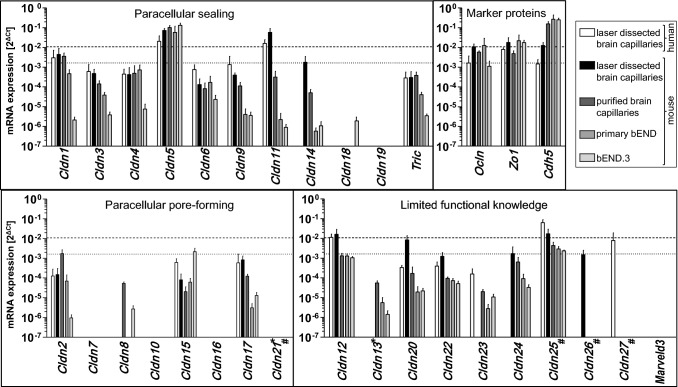
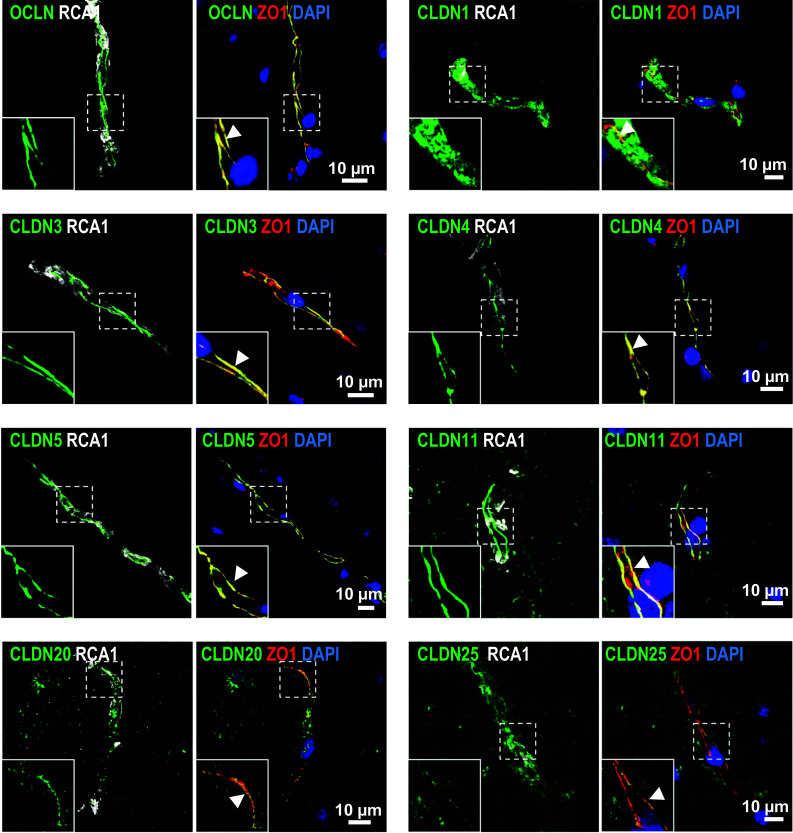
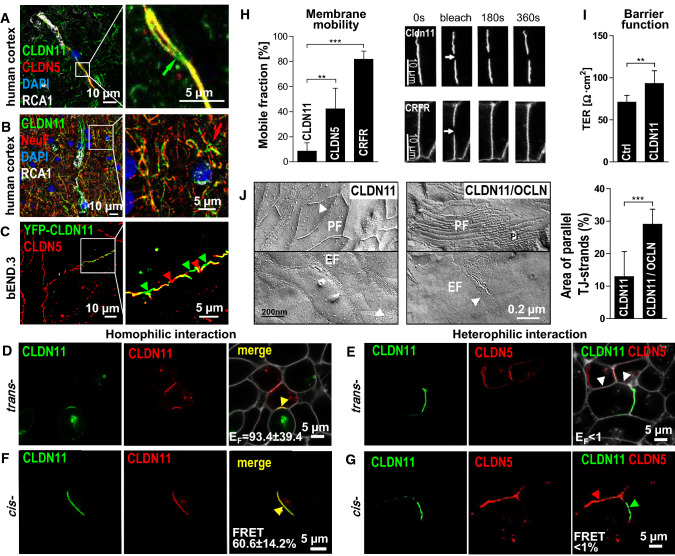
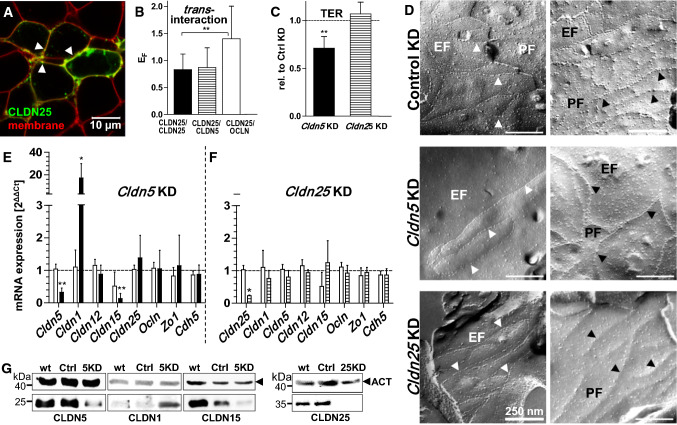
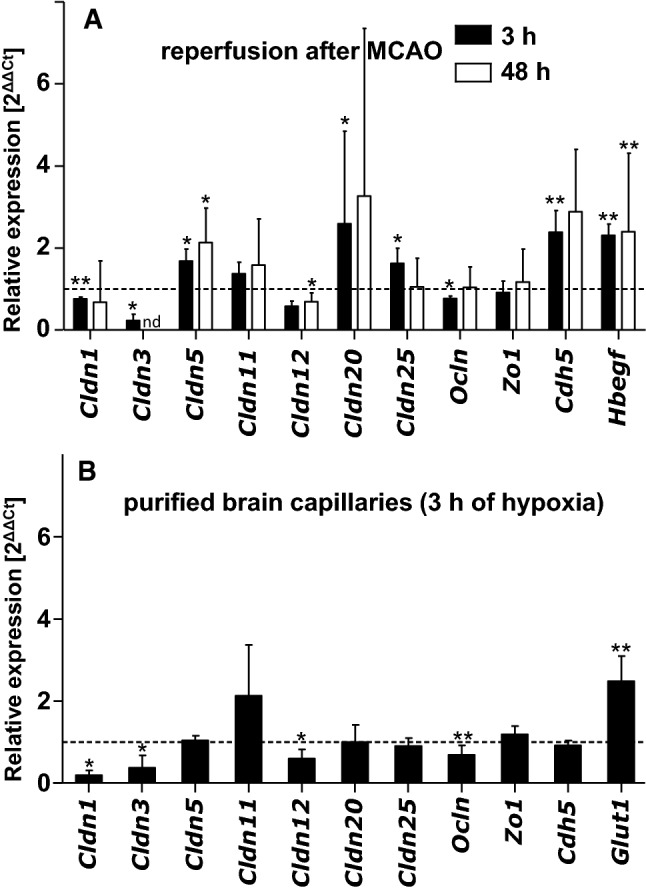
At the blood-brain barrier (BBB), claudin (Cldn)-5 is thought to be the dominant tight junction (TJ) protein, with minor contributions from Cldn3 and -12, and occludin. However, the BBB appears ultrastructurally normal in Cldn5 knock-out mice, suggesting that further Cldns and/or TJ-associated marvel proteins (TAMPs) are involved. Microdissected human and murine brain capillaries, quickly frozen to recapitulate the in vivo situation, showed high transcript expression of Cldn5, -11, -12, and -25, and occludin, but also abundant levels of Cldn1 and -27 in man. Protein levels were quantified by a novel epitope dilution assay and confirmed the respective mRNA data. In contrast to the in vivo situation, Cldn5 dominates BBB expression in vitro, since all other TJ proteins are at comparably low levels or are not expressed. Cldn11 was highly abundant in vivo and contributed to paracellular tightness by homophilic oligomerization, but almost disappeared in vitro. Cldn25, also found at high levels, neither tightened the paracellular barrier nor interconnected opposing cells, but contributed to proper TJ strand morphology. Pathological conditions (in vivo ischemia and in vitro hypoxia) down-regulated Cldn1, -3, and -12, and occludin in cerebral capillaries, which was paralleled by up-regulation of Cldn5 after middle cerebral artery occlusion in rats. Cldn1 expression increased after Cldn5 knock-down. In conclusion, this complete Cldn/TAMP profile demonstrates the presence of up to a dozen TJ proteins in brain capillaries. Mouse and human share a similar and complex TJ profile in vivo, but this complexity is widely lost under in vitro conditions.
Keywords: Brain endothelium; Ischemia; Laser capture microdissection; Neurovasculature; Protein–protein interaction.
Figures

References
-
- Maher GJ, Hilton EN, Urquhart JE, Davidson AE, Spencer HL, Black GC, Manson FD. The cataract-associated protein TMEM114, and TMEM235, are glycosylated transmembrane proteins that are distinct from claudin family members. FEBS Lett. 2011;585(14):2187–2192. doi: 10.1016/j.febslet.2011.05.060. - DOI - PubMed
-
- Cording J, Berg J, Kading N, Bellmann C, Tscheik C, Westphal JK, Milatz S, Gunzel D, Wolburg H, Piontek J, Huber O, Blasig IE. In tight junctions, claudins regulate the interactions between occludin, tricellulin and marvelD3, which, inversely, modulate claudin oligomerization. J Cell Sci. 2013;126(Pt 2):554–564. doi: 10.1242/jcs.114306. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
