

Intratumoural Heterogeneity Underlies Distinct Therapy Responses and Treatment Resistance in Glioblastoma
- PMID: 30736342
- PMCID: PMC6406894
- DOI: 10.3390/cancers11020190
Intratumoural Heterogeneity Underlies Distinct Therapy Responses and Treatment Resistance in Glioblastoma
Abstract
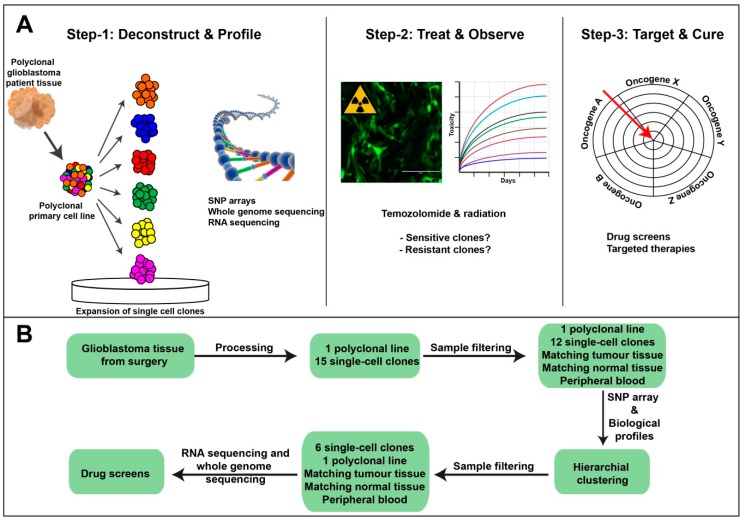
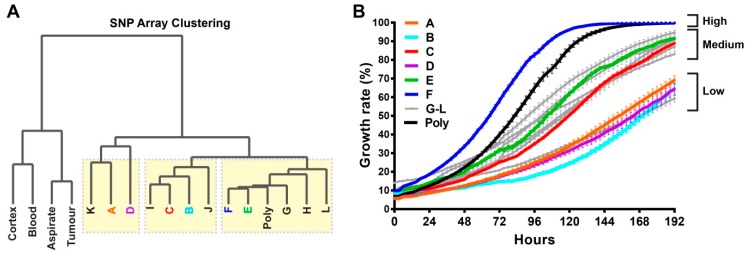
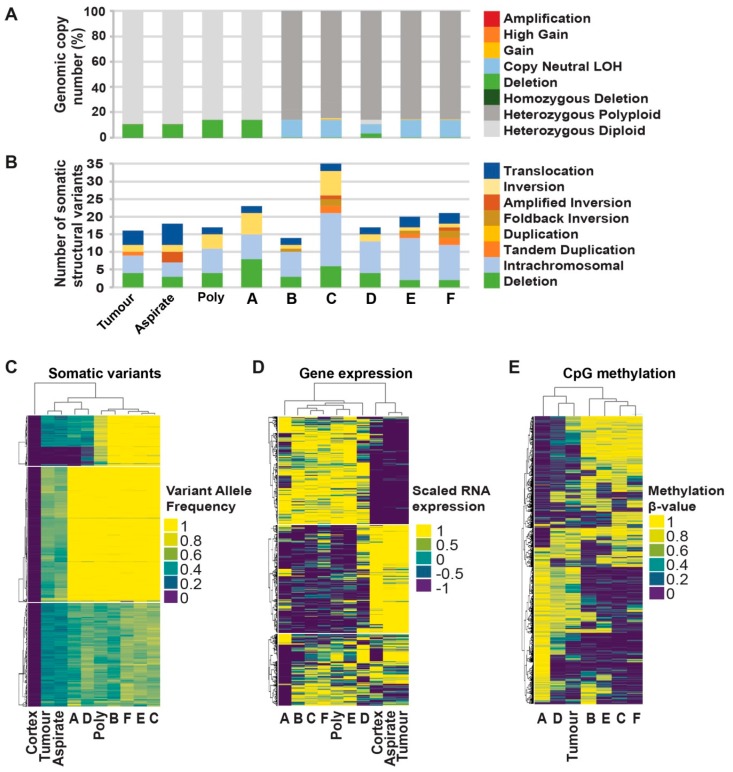
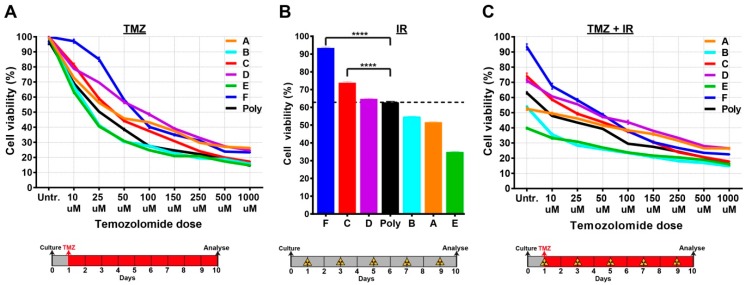
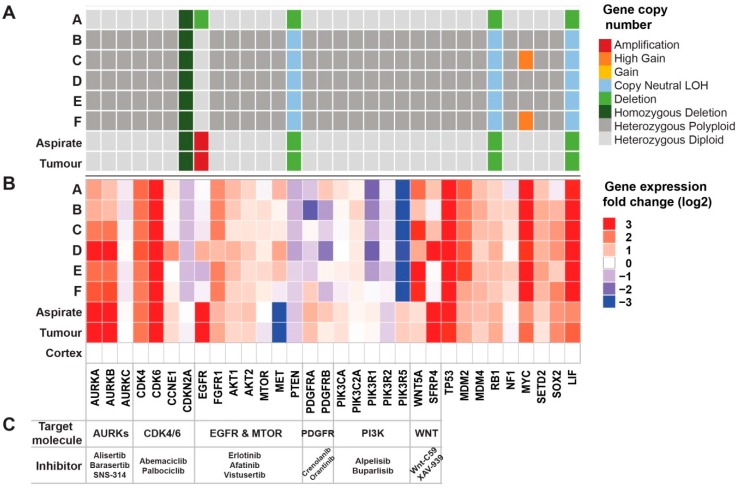
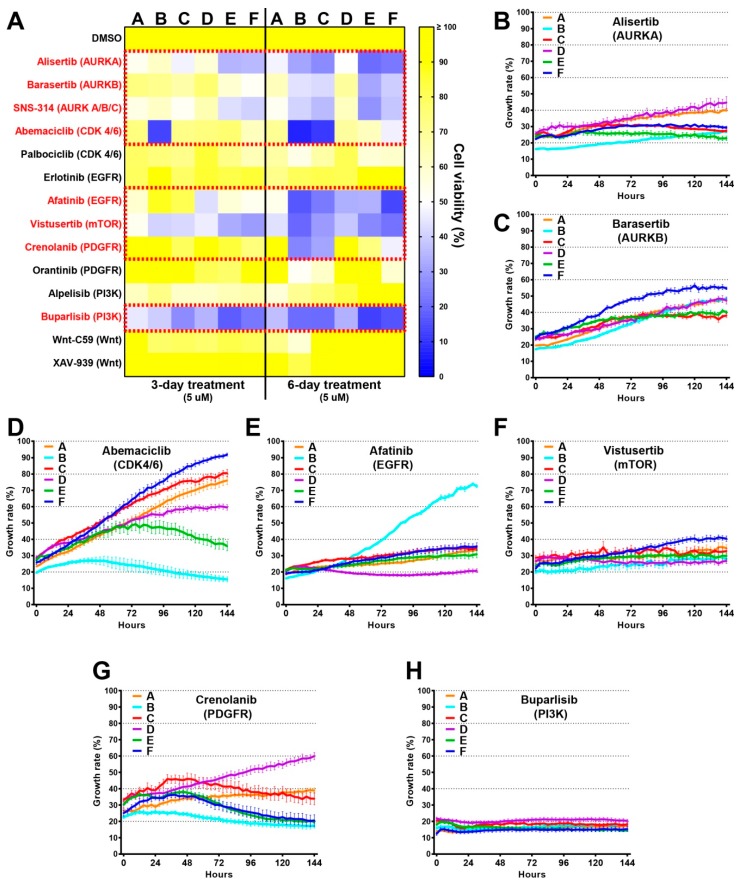
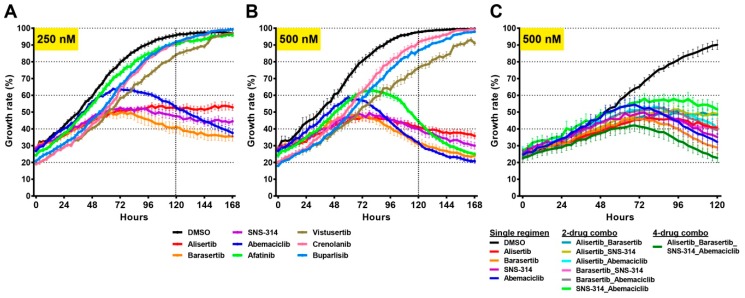
Glioblastomas are the most common and lethal neoplasms of the central nervous system. Neighbouring glioma cells maintain extreme degrees of genetic and phenotypic variation that form intratumoural heterogeneity. This genetic diversity allows the most adaptive tumour clones to develop treatment resistance, ultimately leading to disease recurrence. We aimed to model this phenomenon and test the effectiveness of several targeted therapeutic interventions to overcome therapy resistance. Heterogeneous tumour masses were first deconstructed into single tumour cells, which were expanded independently as single-cell clones. Single nucleotide polymorphism arrays, whole-genome and RNA sequencing, and CpG methylation analysis validated the unique molecular profile of each tumour clone, which displayed distinct pathologic features, including cell morphology, growth rate, and resistance to temozolomide and ionizing radiation. We also identified variable sensitivities to AURK, CDK, and EGFR inhibitors which were consistent with the heterogeneous molecular alterations that each clone harboured. These targeted therapies effectively eliminated the temozolomide- and/or irradiation-resistant clones and also parental polyclonal cells. Our findings indicate that polyclonal tumours create a dynamic environment that consists of diverse tumour elements and treatment responses. Designing targeted therapies based on a range of molecular profiles can be a more effective strategy to eradicate treatment resistance, recurrence, and metastasis.
Keywords: combination therapy; drug screens; glioblastoma; intratumoural heterogeneity; personalised therapy; targeted therapy; tumour resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Verhaak R.G., Hoadley K.A., Purdom E., Wang V., Qi Y., Wilkerson M.D., Miller C.R., Ding L., Golub T., Mesirov J.P., et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell. 2010;17:98–110. doi: 10.1016/j.ccr.2009.12.020. - DOI - PMC - PubMed
-
- Noushmehr H., Weisenberger D.J., Diefes K., Phillips H.S., Pujara K., Berman B.P., Pan F., Pelloski C.E., Sulman E.P., Bhat K.P., et al. Identification of a CpG island methylator phenotype that defines a distinct subgroup of glioma. Cancer Cell. 2010;17:510–522. doi: 10.1016/j.ccr.2010.03.017. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
