

Phosphatases in Mitosis: Roles and Regulation
- PMID: 30736436
- PMCID: PMC6406801
- DOI: 10.3390/biom9020055
Phosphatases in Mitosis: Roles and Regulation
Abstract
Mitosis requires extensive rearrangement of cellular architecture and of subcellular structures so that replicated chromosomes can bind correctly to spindle microtubules and segregate towards opposite poles. This process originates two new daughter nuclei with equal genetic content and relies on highly-dynamic and tightly regulated phosphorylation of numerous cell cycle proteins. A burst in protein phosphorylation orchestrated by several conserved kinases occurs as cells go into and progress through mitosis. The opposing dephosphorylation events are catalyzed by a small set of protein phosphatases, whose importance for the accuracy of mitosis is becoming increasingly appreciated. This review will focus on the established and emerging roles of mitotic phosphatases, describe their structural and biochemical properties, and discuss recent advances in understanding the regulation of phosphatase activity and function.
Keywords: cell division cycle 25 (CDC25); chromosomes; kinetochores; microtubules; mitosis; phosphatases; protein phosphatase 1 (PP1); protein phosphatase 2A (PP2A).
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
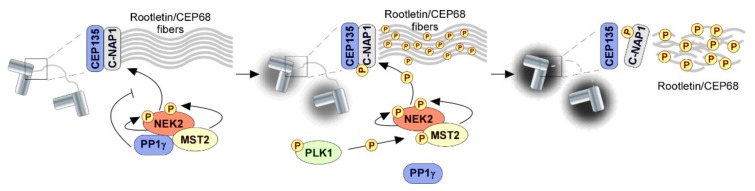
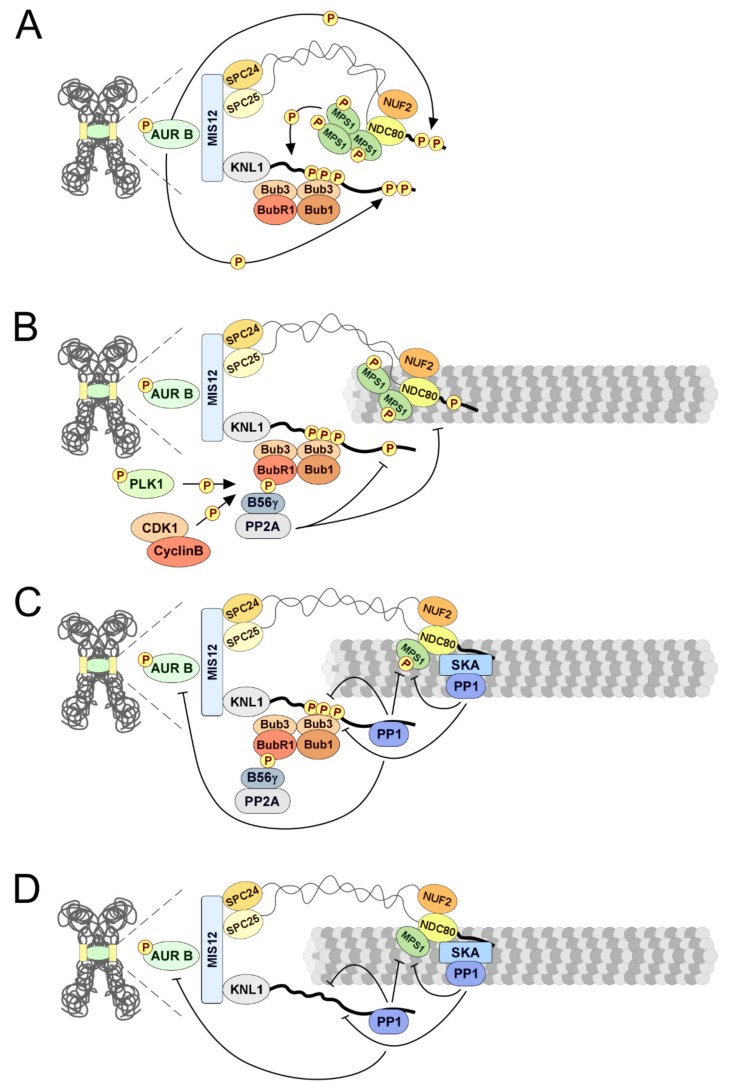
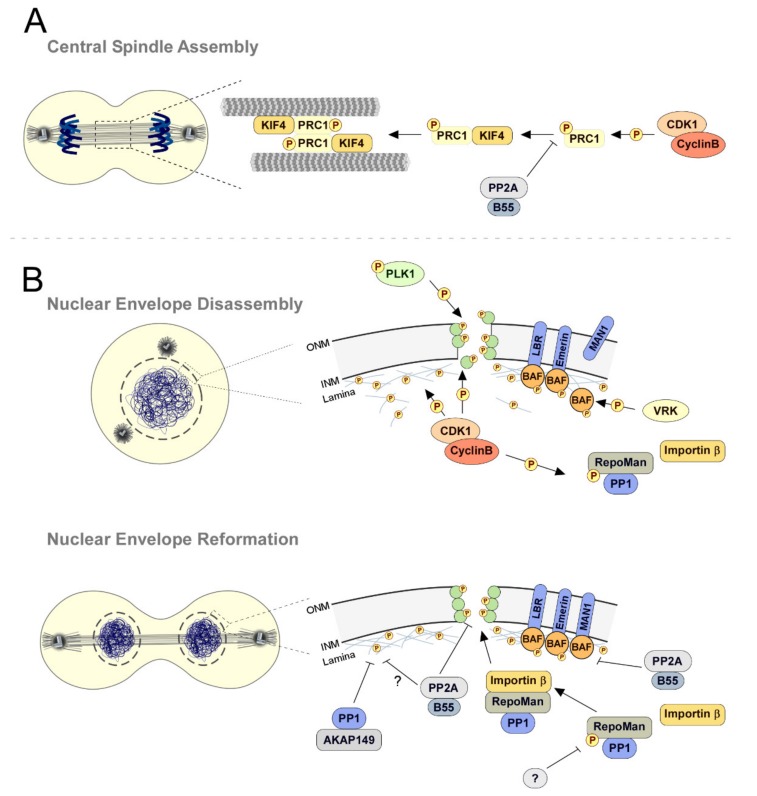
References
-
- Andersen J.N., Mortensen O.H., Peters G.H., Drake P.G., Iversen L.F., Olsen O.H., Jansen P.G., Andersen H.S., Tonks N.K., Møller N.P. Structural and evolutionary relationships among protein tyrosine phosphatase domains. Mol. Cell. Biol. 2001;21:7117–7136. doi: 10.1128/MCB.21.21.7117-7136.2001. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
