

Predicting Polygenic Risk of Psychiatric Disorders
- PMID: 30737014
- PMCID: PMC6599546
- DOI: 10.1016/j.biopsych.2018.12.015
Predicting Polygenic Risk of Psychiatric Disorders
Abstract
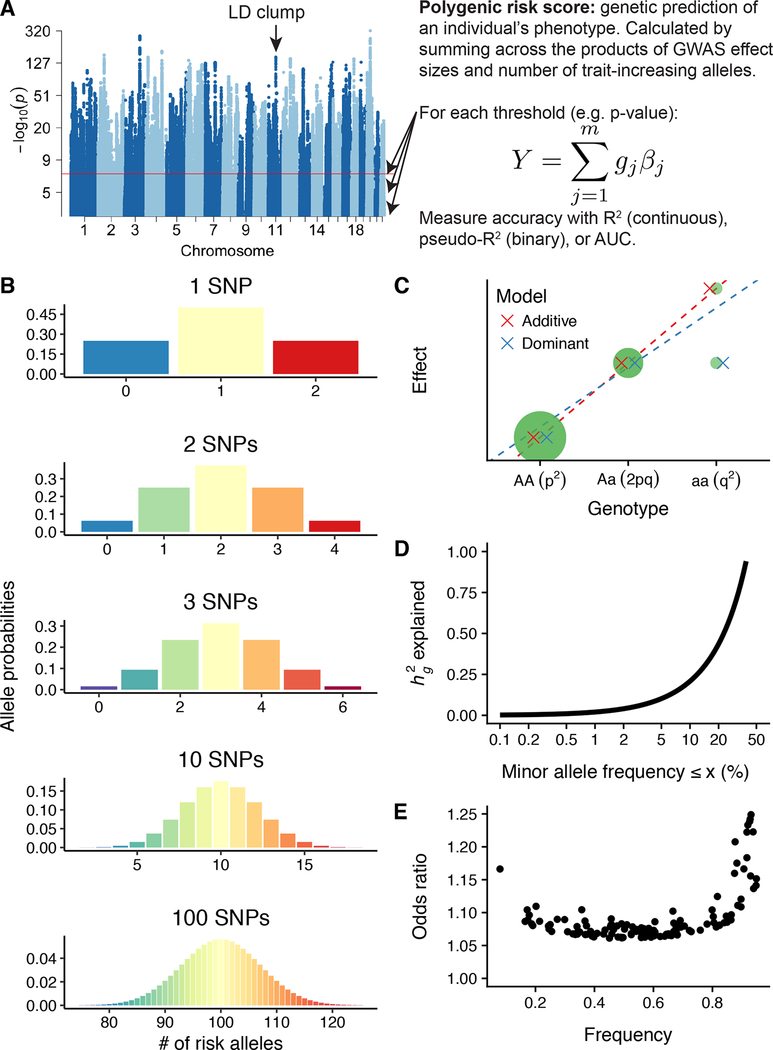
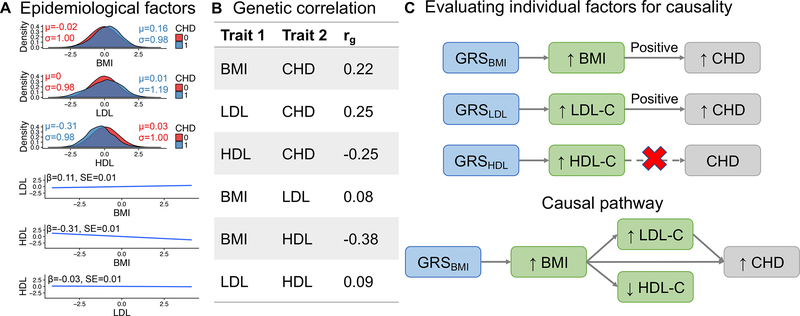
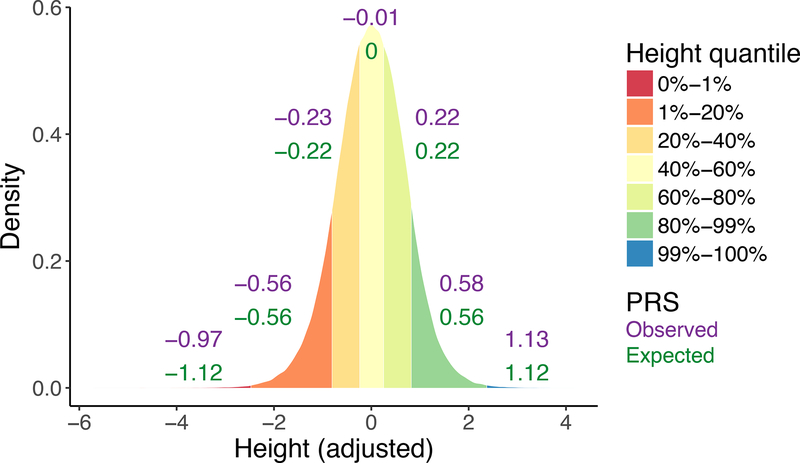
Genetics provides two major opportunities for understanding human disease-as a transformative line of etiological inquiry and as a biomarker for heritable diseases. In psychiatry, biomarkers are very much needed for both research and treatment, given the heterogenous populations identified by current phenomenologically based diagnostic systems. To date, however, useful and valid biomarkers have been scant owing to the inaccessibility and complexity of human brain tissue and consequent lack of insight into disease mechanisms. Genetic biomarkers are therefore especially promising for psychiatric disorders. Genome-wide association studies of common diseases have matured over the last decade, generating the knowledge base for increasingly informative individual-level genetic risk prediction. In this review, we discuss fundamental concepts involved in computing genetic risk with current methods, strengths and weaknesses of various approaches, assessments of utility, and applications to various psychiatric disorders and related traits. Although genetic risk prediction has become increasingly straightforward to apply and common in published studies, there are important pitfalls to avoid. At present, the clinical utility of genetic risk prediction is still low; however, there is significant promise for future clinical applications as the ancestral diversity and sample sizes of genome-wide association studies increase. We discuss emerging data and methods aimed at improving the value of genetic risk prediction for disentangling disease mechanisms and stratifying subjects for epidemiological and clinical studies. For all applications, it is absolutely critical that polygenic risk prediction is applied with appropriate methodology and control for confounding to avoid repeating some mistakes of the candidate gene era.
Keywords: Complex traits; Heritability; Liability threshold model; Polygenic risk scores; Population genetics; Precision medicine; Psychiatric disorders; Psychiatric genetics; Statistical genetics.
Copyright © 2019 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Fisher RA (1918): XV.—The Correlation between Relatives on the Supposition of Mendelian Inheritance. Transactions of the Royal Society of Edinburgh. 52: 399–433.
-
- Pearson K, Lee A (1900): Mathematical Contributions to the Theory of Evolution. VIII. On the Inheritance of Characters not Capable of Exact Quantitative Measurement. Part I. Introductory. Part II. On the Inheritance of Coat-Colour in Horses. Part III. On the Inheritance of Eye-Colour in Man. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences. 195: 79–150.
-
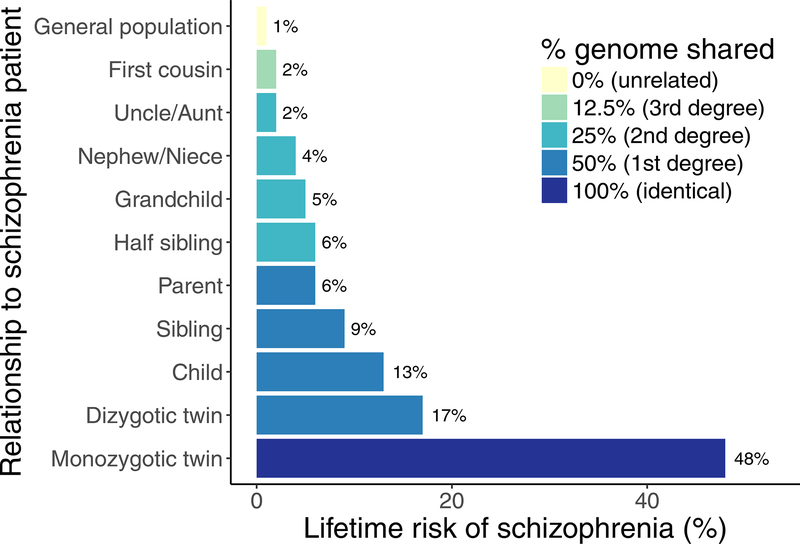
- Falconer DS (1965): The inheritance of liability to certain diseases, estimated from the incidence among relatives. Annals of Human Genetics, 2nd ed. 29: 51–76.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
