

Diet modulates colonic T cell responses by regulating the expression of a Bacteroides thetaiotaomicron antigen
- PMID: 30737355
- PMCID: PMC6550999
- DOI: 10.1126/sciimmunol.aau9079
Diet modulates colonic T cell responses by regulating the expression of a Bacteroides thetaiotaomicron antigen
Abstract
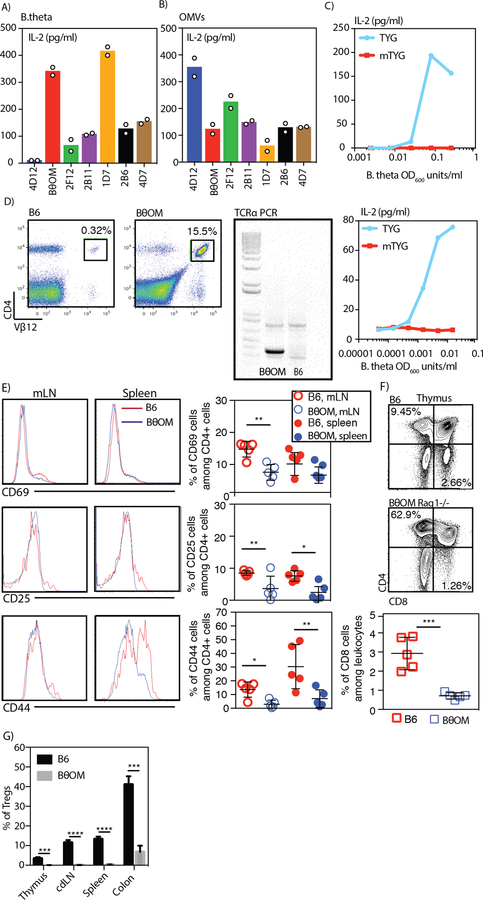
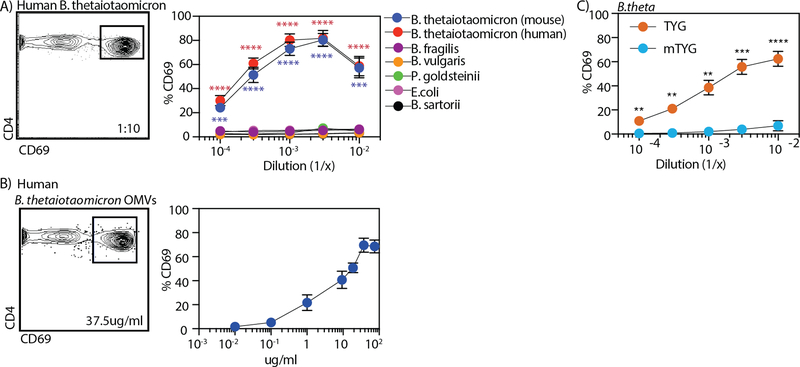
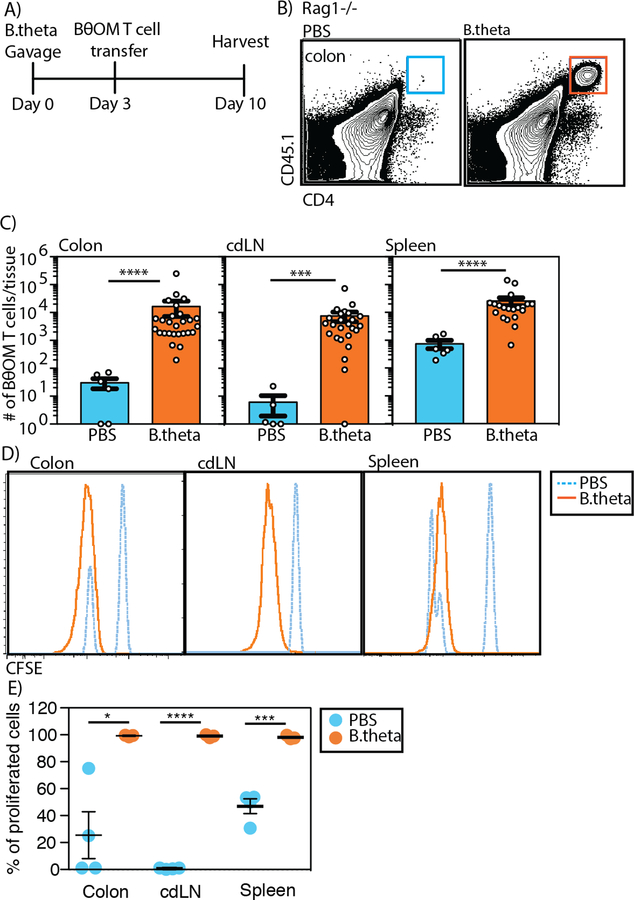
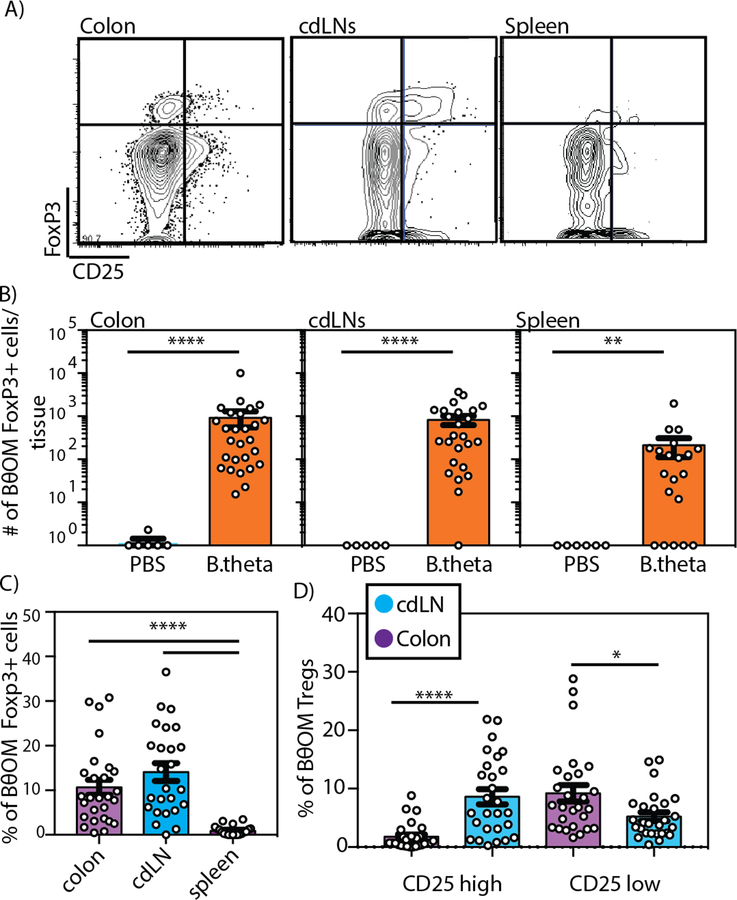
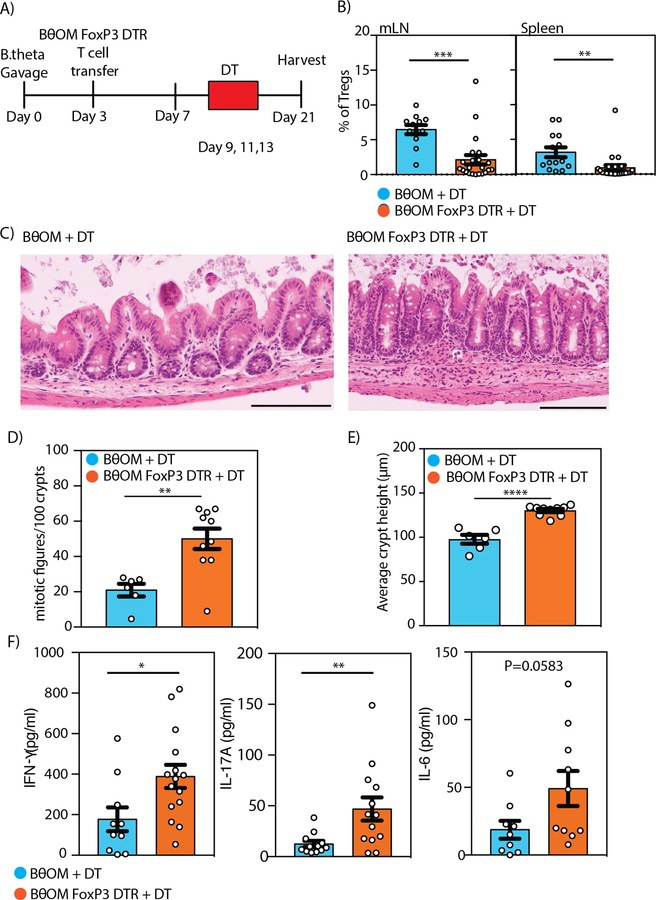
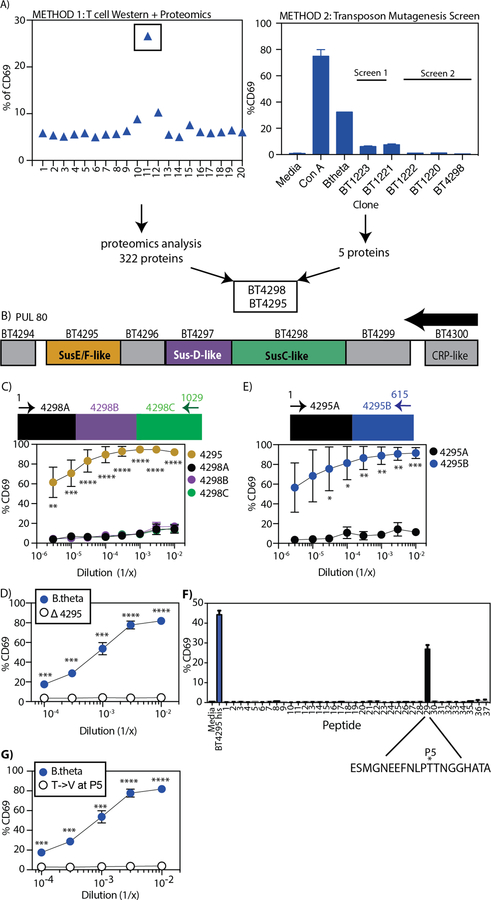
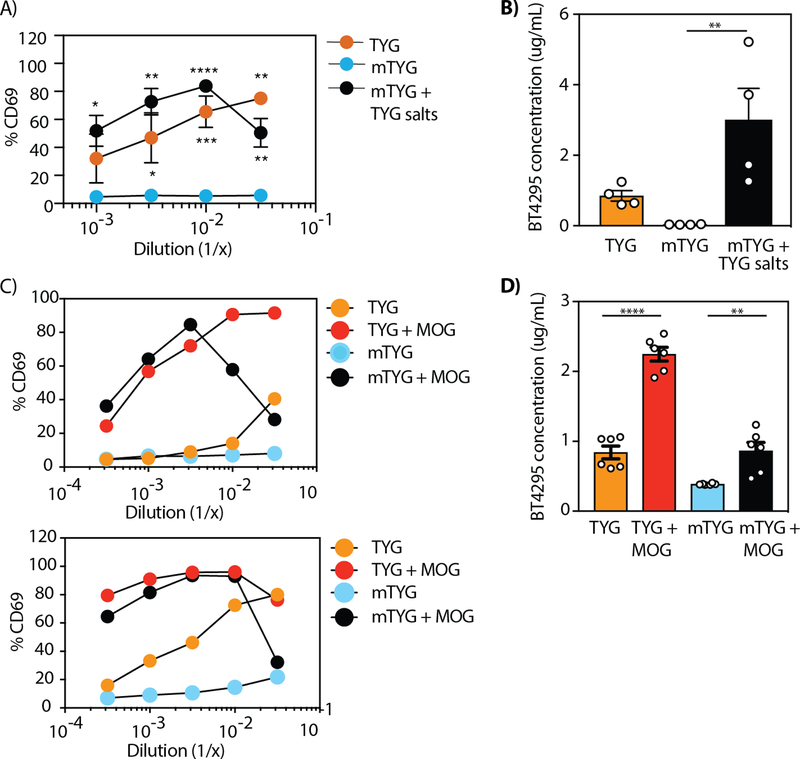
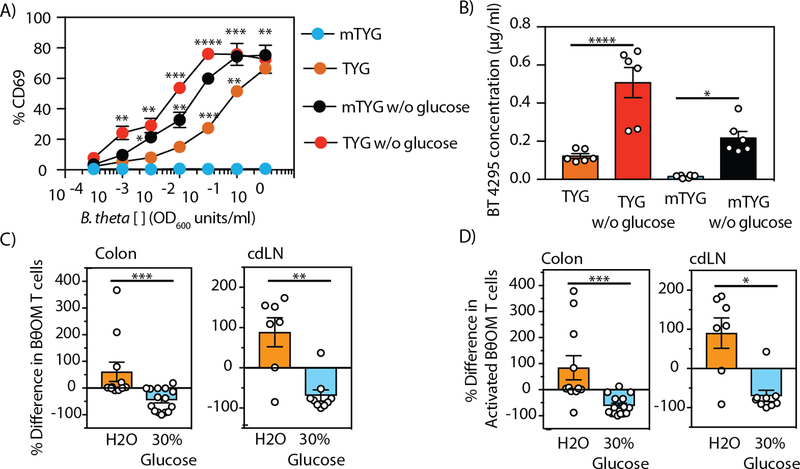
T cell responses to symbionts in the intestine drive tolerance or inflammation depending on the genetic background of the host. These symbionts in the gut sense the available nutrients and adapt their metabolic programs to use these nutrients efficiently. Here, we ask whether diet can alter the expression of a bacterial antigen to modulate adaptive immune responses. We generated a CD4+ T cell hybridoma, BθOM, specific for Bacteroides thetaiotaomicron (B. theta). Adoptively transferred transgenic T cells expressing the BθOM TCR proliferated in the colon, colon-draining lymph node, and spleen in B. theta-colonized healthy mice and differentiated into regulatory T cells (Tregs) and effector T cells (Teffs). Depletion of B. theta-specific Tregs resulted in colitis, showing that a single protein expressed by B. theta can drive differentiation of Tregs that self-regulate Teffs to prevent disease. We found that BθOM T cells recognized a peptide derived from a single B. theta protein, BT4295, whose expression is regulated by nutrients, with glucose being a strong catabolite repressor. Mice fed a high-glucose diet had a greatly reduced activation of BθOM T cells in the colon. These studies establish that the immune response to specific bacterial antigens can be modified by changes in the diet by altering antigen expression in the microbe.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Diet modulates T cell-induced colitis via microbial antigen expression.Nat Rev Gastroenterol Hepatol. 2019 Apr;16(4):198-199. doi: 10.1038/s41575-019-0127-9. Nat Rev Gastroenterol Hepatol. 2019. PMID: 30837701 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
