

Prefrontal Contributions to Attention and Working Memory
- PMID: 30739308
- PMCID: PMC6689265
- DOI: 10.1007/7854_2018_74
Prefrontal Contributions to Attention and Working Memory
Abstract
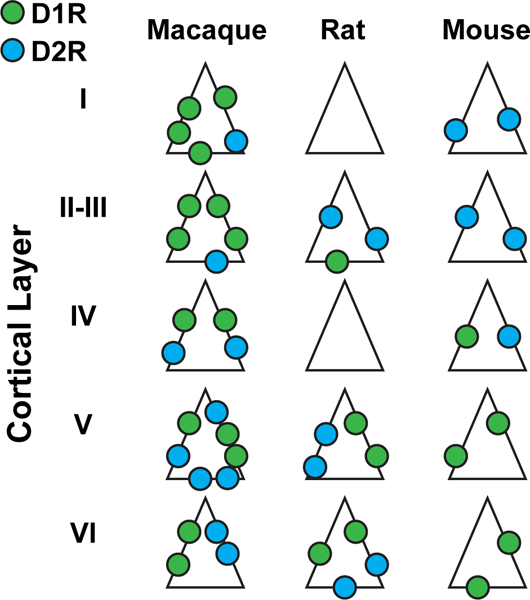
The processes of attention and working memory are conspicuously interlinked, suggesting that they may involve overlapping neural mechanisms. Working memory (WM) is the ability to maintain information in the absence of sensory input. Attention is the process by which a specific target is selected for further processing, and neural resources directed toward that target. The content of WM can be used to direct attention, and attention can in turn determine which information is encoded into WM. Here we discuss the similarities between attention and WM and the role prefrontal cortex (PFC) plays in each. First, at the theoretical level, we describe how attention and WM can both rely on models based on attractor states. Then we review the evidence for an overlap between the areas involved in both functions, especially the frontal eye field (FEF) portion of the prefrontal cortex. We also discuss similarities between the neural changes in visual areas observed during attention and WM. At the cellular level, we review the literature on the role of prefrontal DA in both attention and WM at the behavioral and neural levels. Finally, we summarize the anatomical evidence for an overlap between prefrontal mechanisms involved in attention and WM. Altogether, a summary of pharmacological, electrophysiological, behavioral, and anatomical evidence for a contribution of the FEF part of prefrontal cortex to attention and WM is provided.
Keywords: Attention; Dopamine; Working memory.
Figures
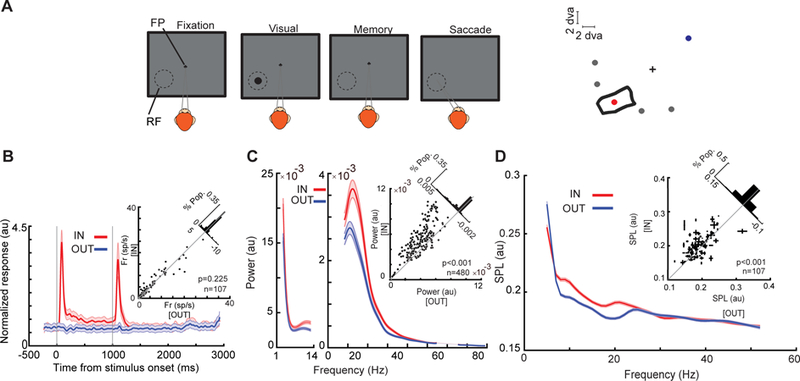
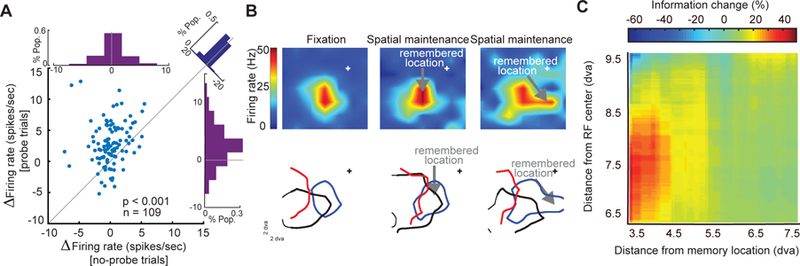
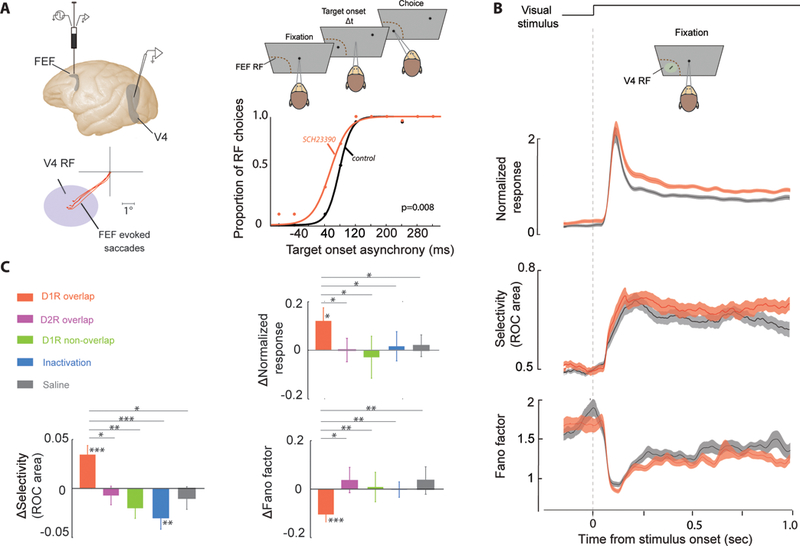
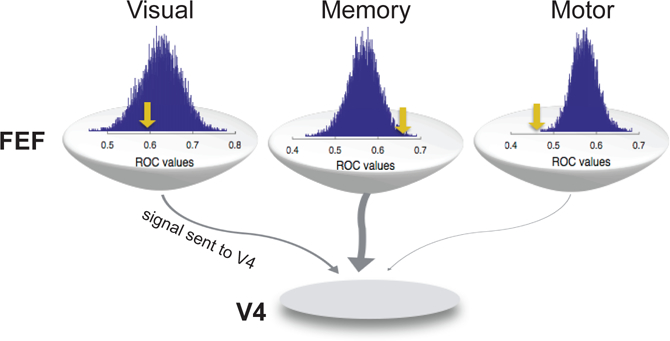
References
-
- Albrecht DG (1995) Visual cortex neurons in monkey and cat: effect of contrast on the spatial and temporal phase transfer functions. Vis Neurosci 12(6):1191–1210Google Scholar - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
