

Formation of Large Native Sulfur Deposits Does Not Require Molecular Oxygen
- PMID: 30740094
- PMCID: PMC6355691
- DOI: 10.3389/fmicb.2019.00024
Formation of Large Native Sulfur Deposits Does Not Require Molecular Oxygen
Abstract
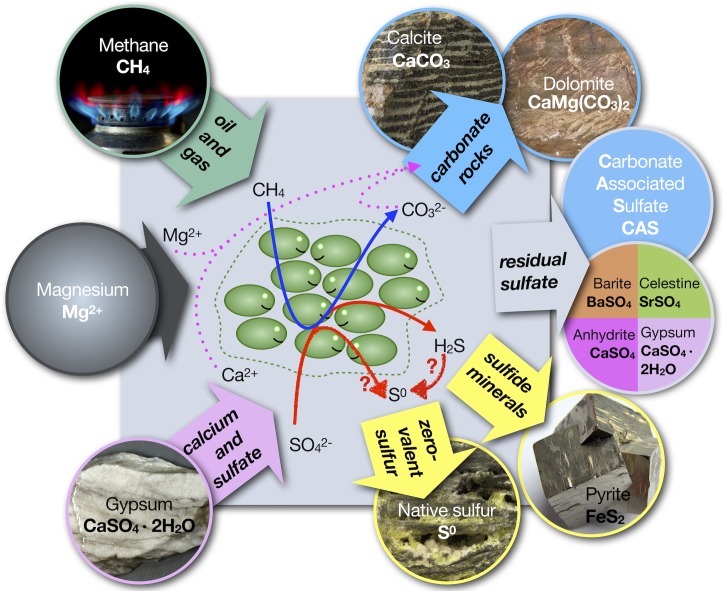
Large native (i.e., elemental) sulfur deposits can be part of caprock assemblages found on top of or in lateral position to salt diapirs and as stratabound mineralization in gypsum and anhydrite lithologies. Native sulfur is formed when hydrocarbons come in contact with sulfate minerals in presence of liquid water. The prevailing model for native sulfur formation in such settings is that sulfide produced by sulfate-reducing bacteria is oxidized to zero-valent sulfur in presence of molecular oxygen (O2). Although possible, such a scenario is problematic because: (1) exposure to oxygen would drastically decrease growth of microbial sulfate-reducing organisms, thereby slowing down sulfide production; (2) on geologic timescales, excess supply with oxygen would convert sulfide into sulfate rather than native sulfur; and (3) to produce large native sulfur deposits, enormous amounts of oxygenated water would need to be brought in close proximity to environments in which ample hydrocarbon supply sustains sulfate reduction. However, sulfur stable isotope data from native sulfur deposits emplaced at a stage after the formation of the host rocks indicate that the sulfur was formed in a setting with little solute exchange with the ambient environment and little supply of dissolved oxygen. We deduce that there must be a process for the formation of native sulfur in absence of an external oxidant for sulfide. We hypothesize that in systems with little solute exchange, sulfate-reducing organisms, possibly in cooperation with other anaerobic microbial partners, drive the formation of native sulfur deposits. In order to cope with sulfide stress, microbes may shift from harmful sulfide production to non-hazardous native sulfur production. We propose four possible mechanisms as a means to form native sulfur: (1) a modified sulfate reduction process that produces sulfur compounds with an intermediate oxidation state, (2) coupling of sulfide oxidation to methanogenesis that utilizes methylated compounds, acetate or carbon dioxide, (3) ammonium oxidation coupled to sulfate reduction, and (4) sulfur comproportionation of sulfate and sulfide. We show these reactions are thermodynamically favorable and especially useful in environments with multiple stressors, such as salt and dissolved sulfide, and provide evidence that microbial species functioning in such environments produce native sulfur. Integrating these insights, we argue that microbes may form large native sulfur deposits in absence of light and external oxidants such as O2, nitrate, and metal oxides. The existence of such a process would not only explain enigmatic occurrences of native sulfur in the geologic record, but also provide an explanation for cryptic sulfur and carbon cycling beneath the seabed.
Keywords: cryptic carbon cycling; cryptic sulfur cycling; isotope; methanogenesis; microbe; native sulfur; sulfur formation; sulfur reduction.
Figures





Similar articles
-
Sulfur and oxygen isotope insights into sulfur cycling in shallow-sea hydrothermal vents, Milos, Greece.Geochem Trans. 2014 Aug 12;15:12. doi: 10.1186/s12932-014-0012-y. eCollection 2014. Geochem Trans. 2014. PMID: 25183951 Free PMC article.
-
Sulfur organic compounds in bottom sediments of the eastern Gulf of Finland.Environ Sci Pollut Res Int. 2007 Sep;14(6):366-76. doi: 10.1065/espr2006.08.334. Environ Sci Pollut Res Int. 2007. PMID: 17993219
-
In situ S-isotope compositions of sulfate and sulfide from the 3.2 Ga Moodies Group, South Africa: A record of oxidative sulfur cycling.Geobiology. 2020 Jul;18(4):426-444. doi: 10.1111/gbi.12393. Epub 2020 Apr 16. Geobiology. 2020. PMID: 32301171
-
The Biogeochemical Sulfur Cycle of Marine Sediments.Front Microbiol. 2019 Apr 24;10:849. doi: 10.3389/fmicb.2019.00849. eCollection 2019. Front Microbiol. 2019. PMID: 31105660 Free PMC article. Review.
-
Dissimilatory reduction of sulfate and zero-valent sulfur at low pH and its significance for bioremediation and metal recovery.Adv Microb Physiol. 2019;75:205-231. doi: 10.1016/bs.ampbs.2019.07.002. Epub 2019 Oct 10. Adv Microb Physiol. 2019. PMID: 31655738 Review.
Cited by
-
Generation of zero-valent sulfur from dissimilatory sulfate reduction in sulfate-reducing microorganisms.Proc Natl Acad Sci U S A. 2023 May 16;120(20):e2220725120. doi: 10.1073/pnas.2220725120. Epub 2023 May 8. Proc Natl Acad Sci U S A. 2023. PMID: 37155857 Free PMC article.
-
An Anaerobic Microbial Community Mediates Epigenetic Native Sulfur and Carbonate Formation During Replacement of Messinian Gypsum at Monte Palco, Sicily.Geobiology. 2025 Mar-Apr;23(2):e70015. doi: 10.1111/gbi.70015. Geobiology. 2025. PMID: 40047358 Free PMC article.
-
Light and polyphosphate kinase 2 cooperatively regulate the production of zero-valent sulfur in a deep-sea bacterium.mSystems. 2025 Jun 17;10(6):e0047325. doi: 10.1128/msystems.00473-25. Epub 2025 May 16. mSystems. 2025. PMID: 40377319 Free PMC article.
-
A novel bacterial thiosulfate oxidation pathway provides a new clue about the formation of zero-valent sulfur in deep sea.ISME J. 2020 Sep;14(9):2261-2274. doi: 10.1038/s41396-020-0684-5. Epub 2020 May 26. ISME J. 2020. PMID: 32457501 Free PMC article.
-
A deep-sea sulfate-reducing bacterium generates zero-valent sulfur via metabolizing thiosulfate.mLife. 2022 Sep 23;1(3):257-271. doi: 10.1002/mlf2.12038. eCollection 2022 Sep. mLife. 2022. PMID: 38818226 Free PMC article.
References
-
- Aeckersberg F., Bak F., Widdel F. (1991). Anaerobic oxidation of saturated hydrocarbons to CO2 by a new type of sulfate-reducing bacterium. Arch. Microbiol. 156 5–14. 10.1007/BF00418180 - DOI
-
- Akagi J. M. (1995). “Respiratory sulfate reduction,” in Sulfate-Reducing Bacteria Biotechnology Handbooks, eds Barton L. L., Sherwood R. F. (New York, NY: Plenum Press; ), 89–111. 10.1007/978-1-4899-1582-5_4 - DOI
-
- Aloisi G., Baudrand M., Lécuyer C., Rouchy J.-M., Pancost R. D., Aref M. A. M., et al. (2013). Biomarker and isotope evidence for microbially-mediated carbonate formation from gypsum and petroleum hydrocarbons. Chem. Geol. 347 199–207. 10.1016/j.chemgeo.2013.03.007 - DOI
LinkOut - more resources
Full Text Sources