

Mosaic nanoparticle display of diverse influenza virus hemagglutinins elicits broad B cell responses
- PMID: 30742080
- PMCID: PMC6380945
- DOI: 10.1038/s41590-018-0305-x
Mosaic nanoparticle display of diverse influenza virus hemagglutinins elicits broad B cell responses
Erratum in
-
Author Correction: Mosaic nanoparticle display of diverse influenza virus hemagglutinins elicits broad B cell responses.Nat Immunol. 2019 Jun;20(6):765. doi: 10.1038/s41590-019-0395-0. Nat Immunol. 2019. PMID: 30980067
Abstract
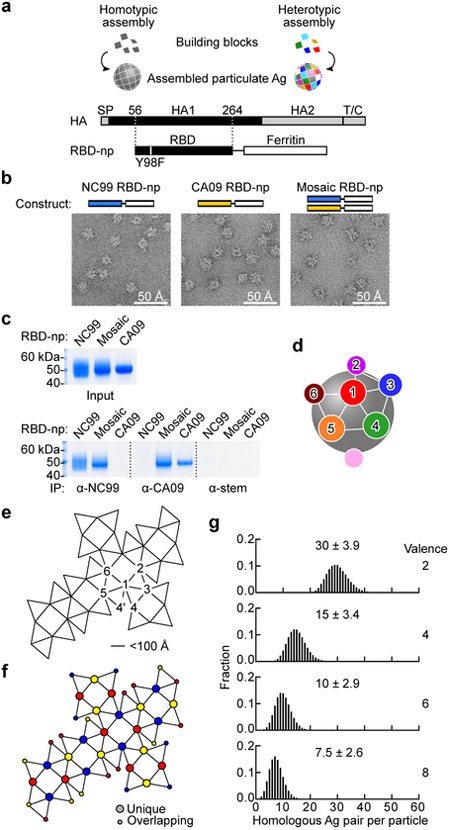
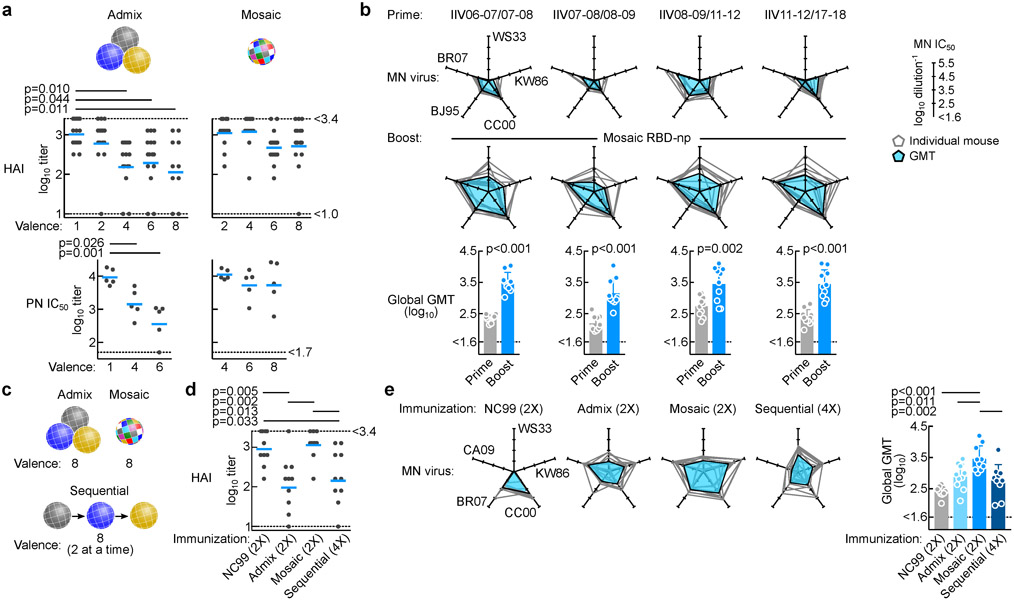
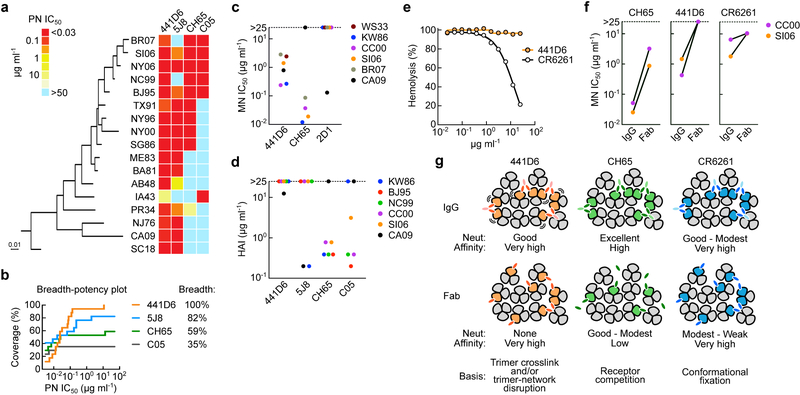
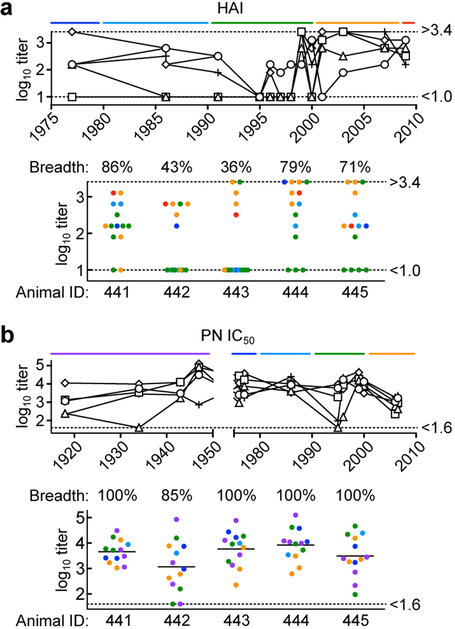
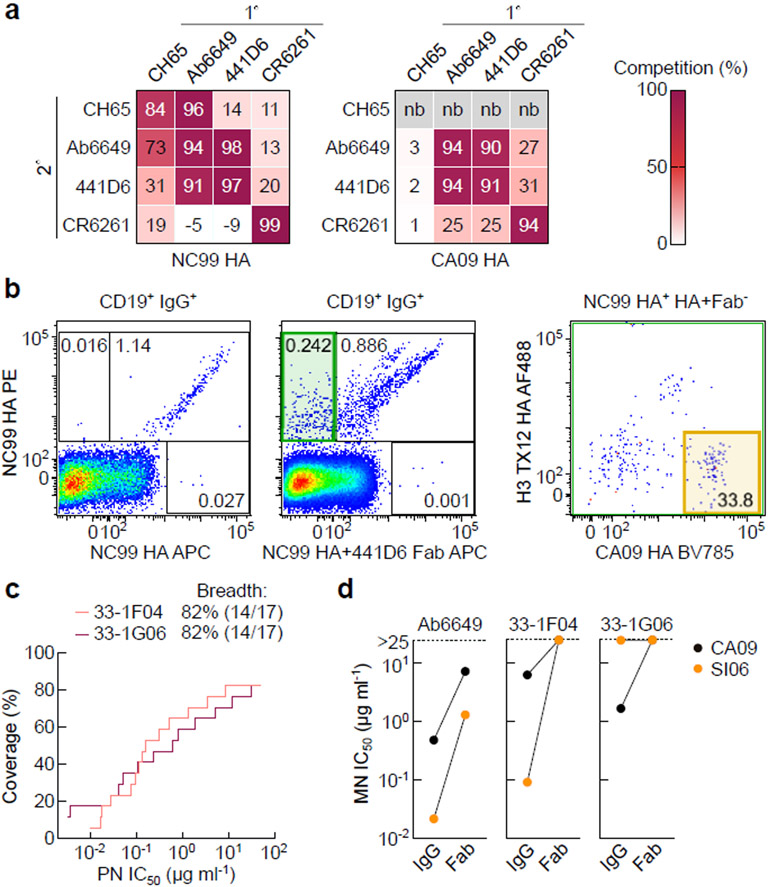
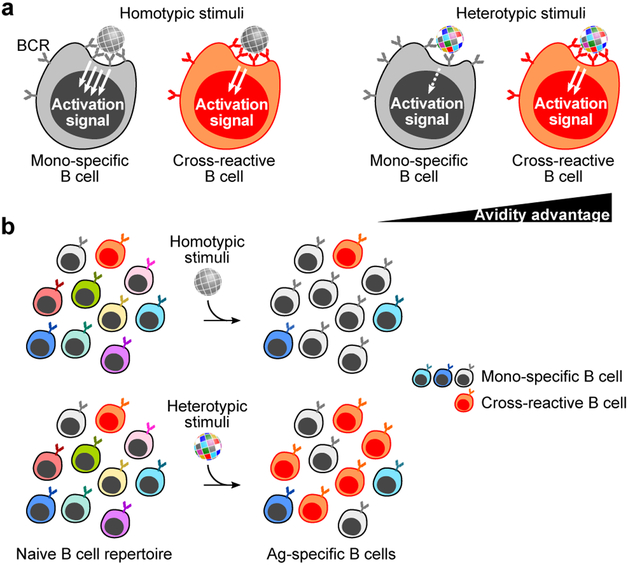
The present vaccine against influenza virus has the inevitable risk of antigenic discordance between the vaccine and the circulating strains, which diminishes vaccine efficacy. This necessitates new approaches that provide broader protection against influenza. Here we designed a vaccine using the hypervariable receptor-binding domain (RBD) of viral hemagglutinin displayed on a nanoparticle (np) able to elicit antibody responses that neutralize H1N1 influenza viruses spanning over 90 years. Co-display of RBDs from multiple strains across time, so that the adjacent RBDs are heterotypic, provides an avidity advantage to cross-reactive B cells. Immunization with the mosaic RBD-np elicited broader antibody responses than those induced by an admixture of nanoparticles encompassing the same set of RBDs as separate homotypic arrays. Furthermore, we identified a broadly neutralizing monoclonal antibody in a mouse immunized with mosaic RBD-np. The mosaic antigen array signifies a unique approach that subverts monotypic immunodominance and allows otherwise subdominant cross-reactive B cell responses to emerge.
Figures
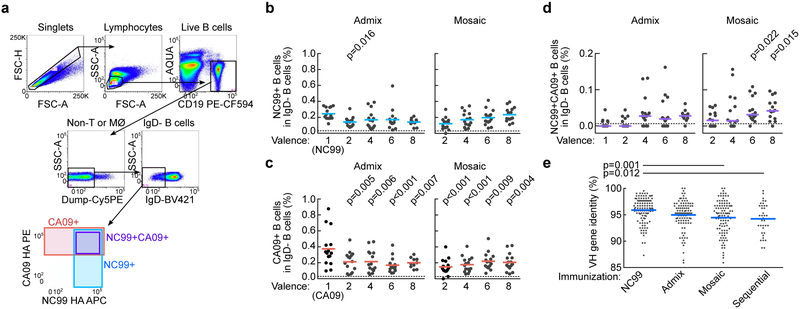
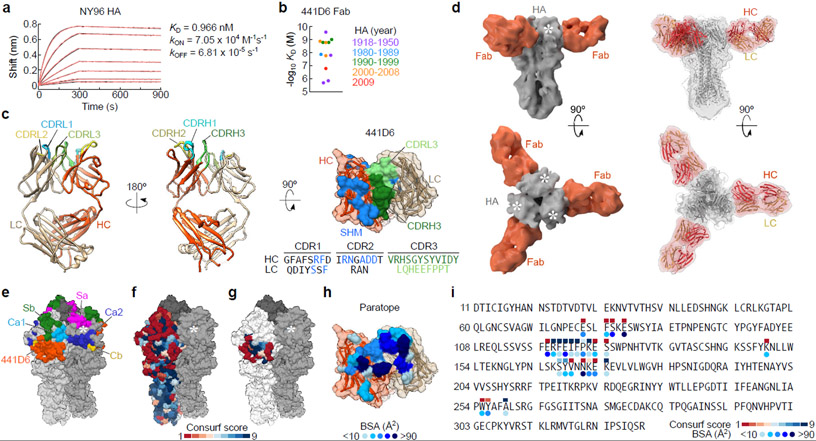
Comment in
-
Fighting influenza through hemagglutinin diversity.Nat Immunol. 2019 Mar;20(3):246-247. doi: 10.1038/s41590-019-0317-1. Nat Immunol. 2019. PMID: 30742079 No abstract available.
References
-
- Koel BF et al. Substitutions near the receptor binding site determine major antigenic change during influenza virus evolution. Science 342, 976–979 (2013). - PubMed
-
- Smith DJ et al. Mapping the antigenic and genetic evolution of influenza virus. Science 305, 371–376 (2004). - PubMed
-
- Webster RG, Laver WG, Air GM & Schild GC Molecular mechanisms of variation in influenza viruses. Nature 296, 115–121 (1982). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
