

Regulation of BK Channels by Beta and Gamma Subunits
- PMID: 30742788
- PMCID: PMC6380188
- DOI: 10.1146/annurev-physiol-022516-034038
Regulation of BK Channels by Beta and Gamma Subunits
Abstract
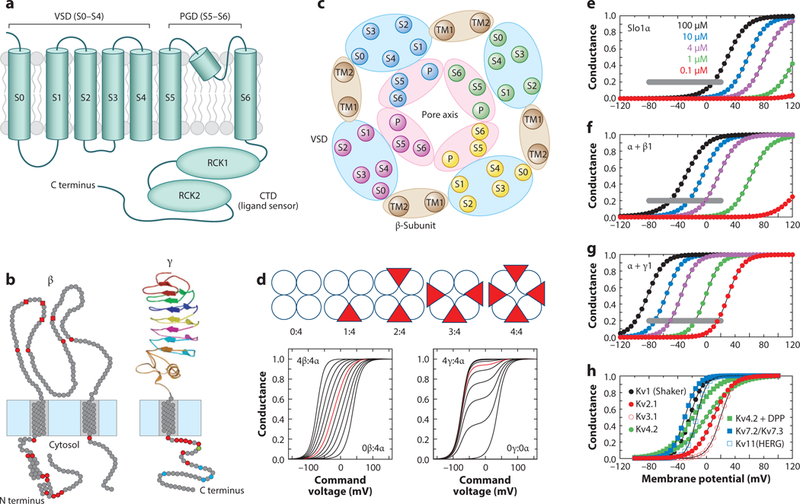
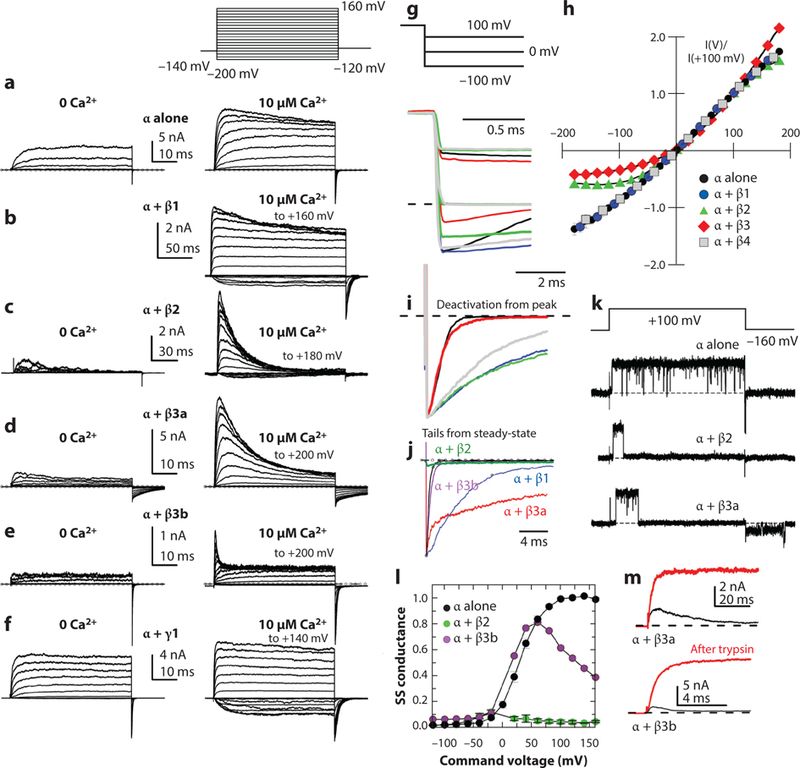
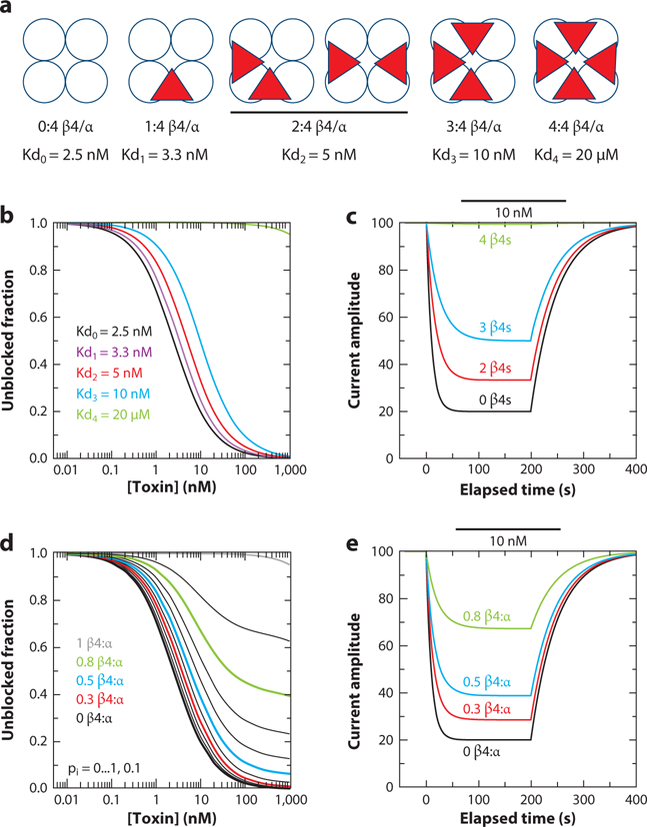
Ca2+- and voltage-gated K+ channels of large conductance (BK channels) are expressed in a diverse variety of both excitable and inexcitable cells, with functional properties presumably uniquely calibrated for the cells in which they are found. Although some diversity in BK channel function, localization, and regulation apparently arises from cell-specific alternative splice variants of the single pore-forming α subunit ( KCa1.1, Kcnma1, Slo1) gene, two families of regulatory subunits, β and γ, define BK channels that span a diverse range of functional properties. We are just beginning to unravel the cell-specific, physiological roles served by BK channels of different subunit composition.
Keywords: BK channels; Ca- and voltage-dependent K channels; auxiliary subunits; beta subunits; gamma subunits.
Figures
Similar articles
-
Regulation of BK channels by auxiliary γ subunits.Front Physiol. 2014 Oct 15;5:401. doi: 10.3389/fphys.2014.00401. eCollection 2014. Front Physiol. 2014. PMID: 25360119 Free PMC article. Review.
-
Dysregulation of Podocyte BK Channels and Nephrosis: Effects of Circulating Factors and Auxiliary β4 Subunits.Cells. 2024 Dec 30;14(1):22. doi: 10.3390/cells14010022. Cells. 2024. PMID: 39791723 Free PMC article.
-
Molecular structure and function of big calcium-activated potassium channels in skeletal muscle: pharmacological perspectives.Physiol Genomics. 2017 Jun 1;49(6):306-317. doi: 10.1152/physiolgenomics.00121.2016. Epub 2017 Apr 28. Physiol Genomics. 2017. PMID: 28455309
-
Regulatory γ1 subunits defy symmetry in functional modulation of BK channels.Proc Natl Acad Sci U S A. 2018 Oct 2;115(40):9923-9928. doi: 10.1073/pnas.1804560115. Epub 2018 Sep 17. Proc Natl Acad Sci U S A. 2018. PMID: 30224470 Free PMC article.
-
Modulation of BK Channel Function by Auxiliary Beta and Gamma Subunits.Int Rev Neurobiol. 2016;128:51-90. doi: 10.1016/bs.irn.2016.03.015. Epub 2016 Apr 8. Int Rev Neurobiol. 2016. PMID: 27238261 Free PMC article. Review.
Cited by
-
Regulation of the Mitochondrial BKCa Channel by the Citrus Flavonoid Naringenin as a Potential Means of Preventing Cell Damage.Molecules. 2020 Jun 30;25(13):3010. doi: 10.3390/molecules25133010. Molecules. 2020. PMID: 32630135 Free PMC article.
-
Kinetics and functional consequences of BK channels activation by N-type Ca2+ channels in the dendrite of mouse neocortical layer-5 pyramidal neurons.Front Cell Neurosci. 2024 Feb 14;18:1353895. doi: 10.3389/fncel.2024.1353895. eCollection 2024. Front Cell Neurosci. 2024. PMID: 38419657 Free PMC article.
-
BKCa Channels as Targets for Cardioprotection.Antioxidants (Basel). 2020 Aug 17;9(8):760. doi: 10.3390/antiox9080760. Antioxidants (Basel). 2020. PMID: 32824463 Free PMC article. Review.
-
A review of the pathophysiology and the role of ion channels on bronchial asthma.Front Pharmacol. 2023 Sep 28;14:1236550. doi: 10.3389/fphar.2023.1236550. eCollection 2023. Front Pharmacol. 2023. PMID: 37841931 Free PMC article. Review.
-
Effect of an autism-associated KCNMB2 variant, G124R, on BK channel properties.Curr Res Physiol. 2022 Sep 25;5:404-413. doi: 10.1016/j.crphys.2022.09.001. eCollection 2022. Curr Res Physiol. 2022. PMID: 36203817 Free PMC article.
References
-
- Adelman JP, Shen KZ, Kavanaugh MP, Warren RA, Wu YN, et al. 1992. Calcium-activated potassium channels expressed from cloned complementary DNAs. Neuron 9:209–16 - PubMed
-
- Butler A, Tsunoda S, McCobb DP, Wei A, Salkoff L. 1993. mSlo, a complex mouse gene encoding “maxi” calcium-activated potassium channels. Science 261:221–24 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
