

Neural Substrates of Drosophila Larval Anemotaxis
- PMID: 30744969
- PMCID: PMC6380933
- DOI: 10.1016/j.cub.2019.01.009
Neural Substrates of Drosophila Larval Anemotaxis
Abstract
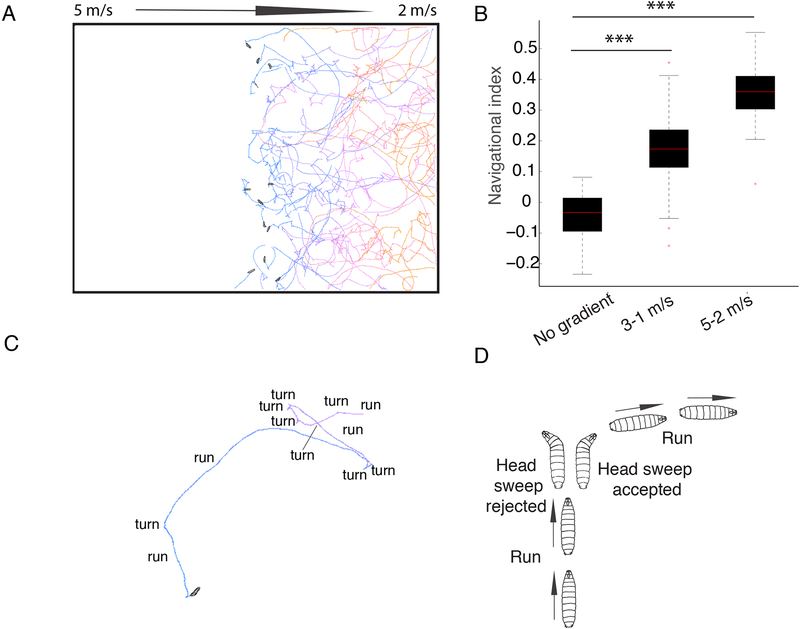
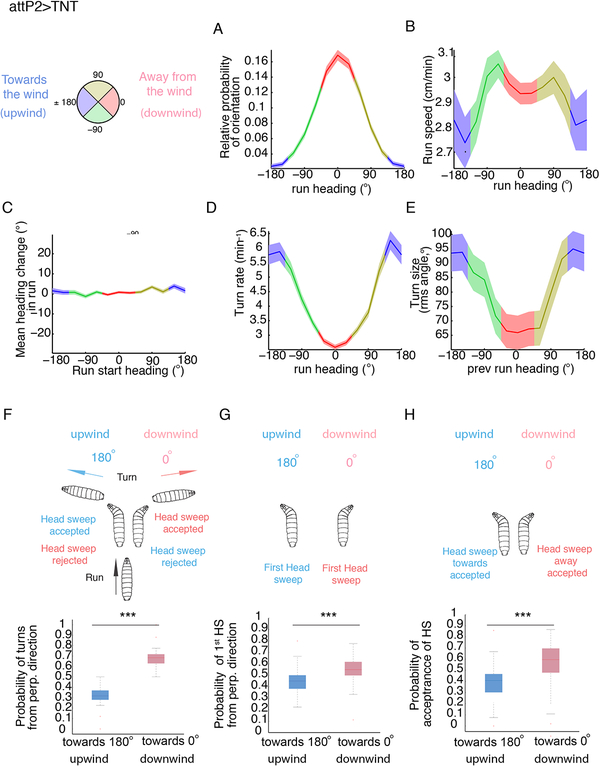
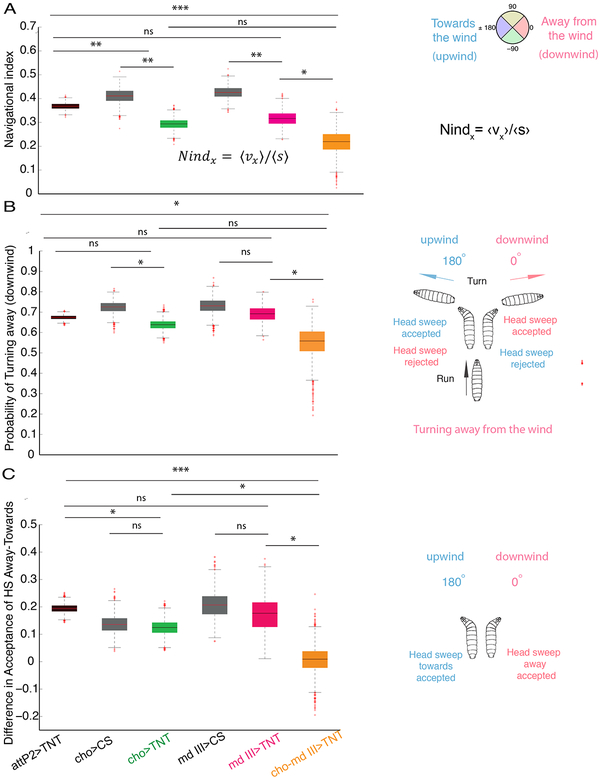
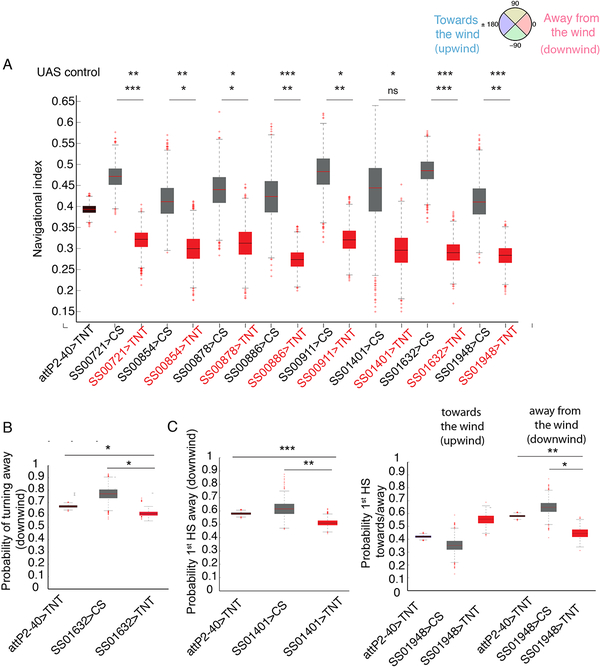
Animals use sensory information to move toward more favorable conditions. Drosophila larvae can move up or down gradients of odors (chemotax), light (phototax), and temperature (thermotax) by modulating the probability, direction, and size of turns based on sensory input. Whether larvae can anemotax in gradients of mechanosensory cues is unknown. Further, although many of the sensory neurons that mediate taxis have been described, the central circuits are not well understood. Here, we used high-throughput, quantitative behavioral assays to demonstrate Drosophila larvae anemotax in gradients of wind speeds and to characterize the behavioral strategies involved. We found that larvae modulate the probability, direction, and size of turns to move away from higher wind speeds. This suggests that similar central decision-making mechanisms underlie taxis in somatosensory and other sensory modalities. By silencing the activity of single or very few neuron types in a behavioral screen, we found two sensory (chordotonal and multidendritic class III) and six nerve cord neuron types involved in anemotaxis. We reconstructed the identified neurons in an electron microscopy volume that spans the entire larval nervous system and found they received direct input from the mechanosensory neurons or from each other. In this way, we identified local interneurons and first- and second-order subesophageal zone (SEZ) and brain projection neurons. Finally, silencing a dopaminergic brain neuron type impairs anemotaxis. These findings suggest that anemotaxis involves both nerve cord and brain circuits. The candidate neurons and circuitry identified in our study provide a basis for future detailed mechanistic understanding of the circuit principles of anemotaxis.
Keywords: CNS; Drosophila larva; anemotaxis; neural substrates; somatosensory processing.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interest.
Figures
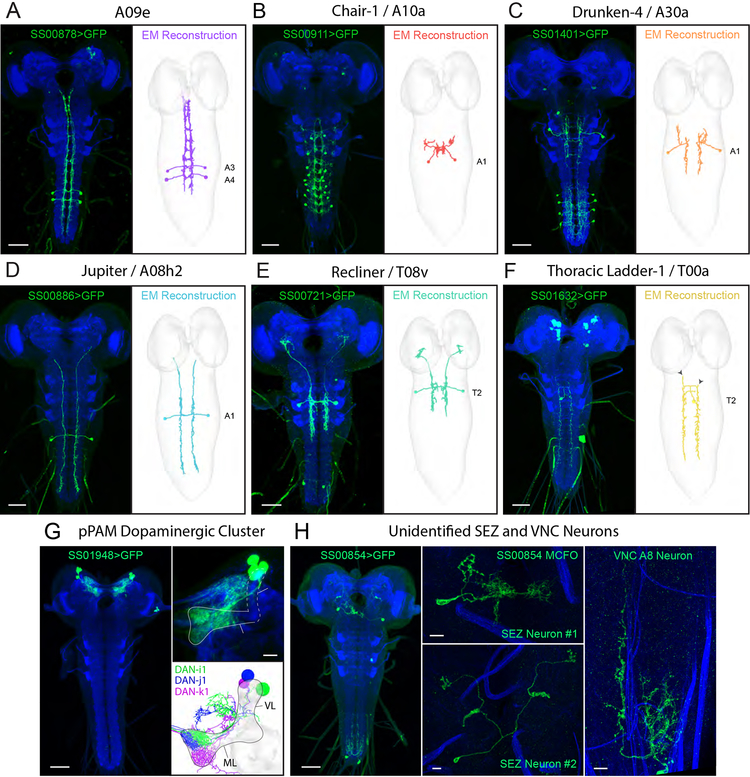
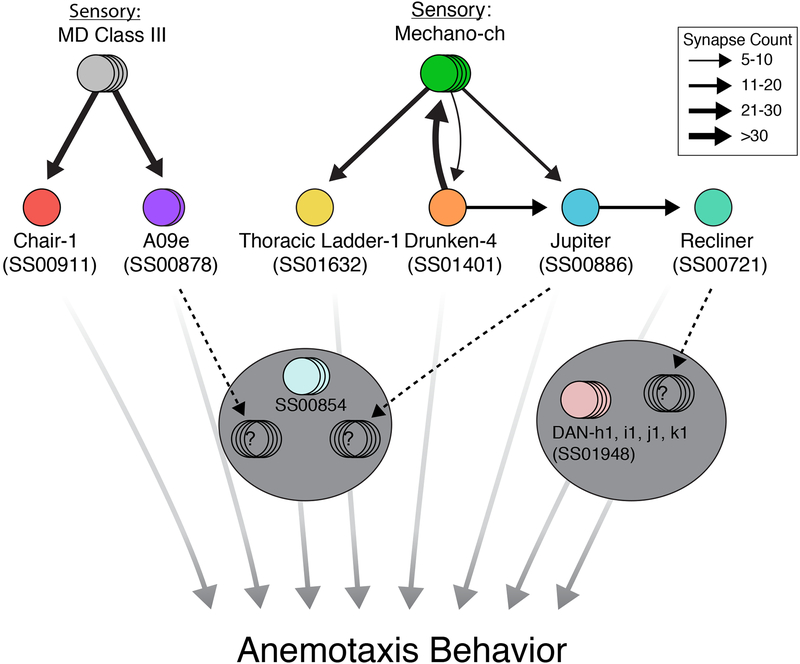
References
-
- Van Tilborg M, Sabelis MW, and Roessingh P (2004). State-dependent and odour-mediated anemotactic responses of the predatory mite Phytoseiulus persimilis in a wind tunnel. Exp. Appl. Acarol 32, 263–270. - PubMed
-
- Kalmus H (1942). Anemotaxis in Drosophila. Nature 150, 405–1.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
