

Haematopoietic stem cell activity and interactions with the niche
- PMID: 30745579
- PMCID: PMC6483843
- DOI: 10.1038/s41580-019-0103-9
Haematopoietic stem cell activity and interactions with the niche
Abstract
The haematopoietic stem cell (HSC) microenvironment in the bone marrow, termed the niche, ensures haematopoietic homeostasis by controlling the proliferation, self-renewal, differentiation and migration of HSCs and progenitor cells at steady state and in response to emergencies and injury. Improved methods for HSC isolation, driven by advances in single-cell and molecular technologies, have led to a better understanding of their behaviour, heterogeneity and lineage fate and of the niche cells and signals that regulate their function. Niche regulatory signals can be in the form of cell-bound or secreted factors and other local physical cues. A combination of technological advances in bone marrow imaging and genetic manipulation of crucial regulatory factors has enabled the identification of several candidate cell types regulating the niche, including both non-haematopoietic (for example, perivascular mesenchymal stem and endothelial cells) and HSC-derived (for example, megakaryocytes, macrophages and regulatory T cells), with better topographical understanding of HSC localization in the bone marrow. Here, we review advances in our understanding of HSC regulation by niches during homeostasis, ageing and cancer, and we discuss their implications for the development of therapies to rejuvenate aged HSCs or niches or to disrupt self-reinforcing malignant niches.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
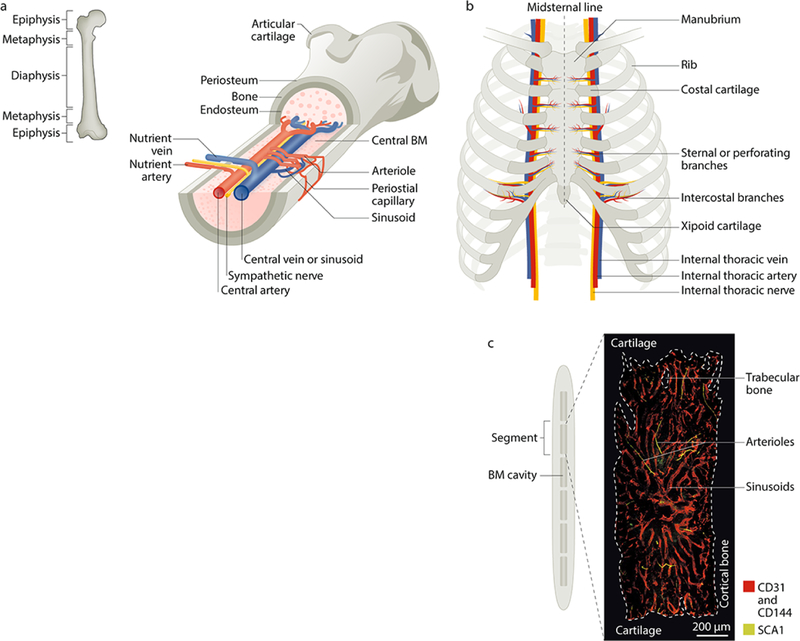

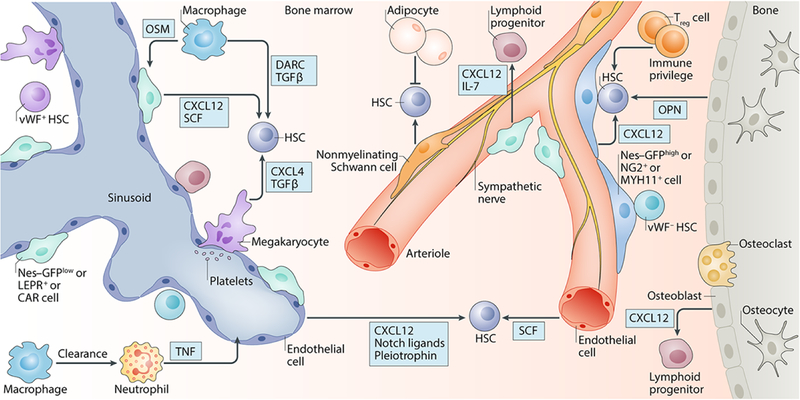

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
