

Immunobiochemical Reconstruction of Influenza Lung Infection-Melanoma Skin Cancer Interactions
- PMID: 30745900
- PMCID: PMC6360404
- DOI: 10.3389/fimmu.2019.00004
Immunobiochemical Reconstruction of Influenza Lung Infection-Melanoma Skin Cancer Interactions
Abstract
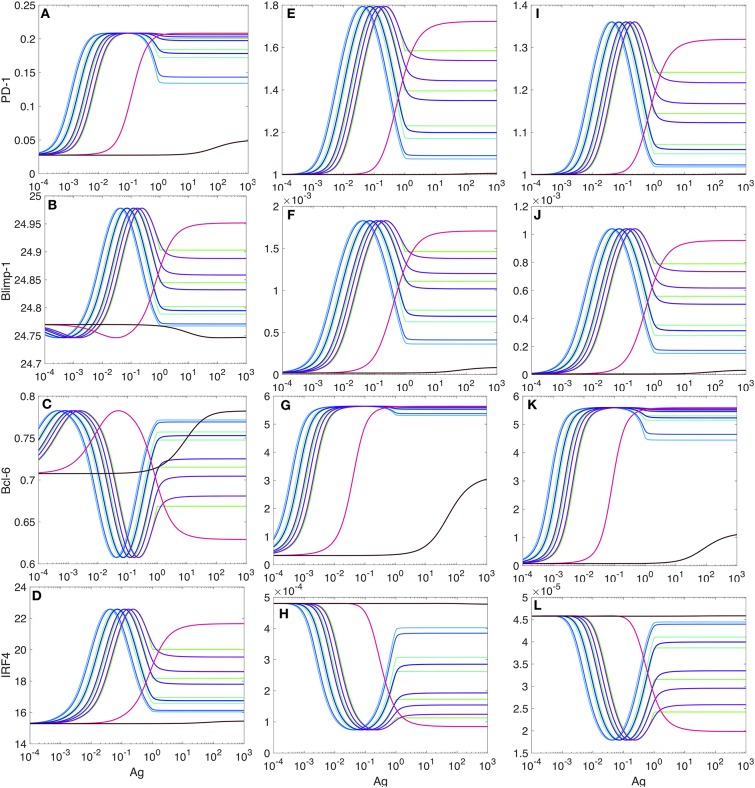
It was recently reported that acute influenza infection of the lung promoted distal melanoma growth in the dermis of mice. Melanoma-specific CD8+ T cells were shunted to the lung in the presence of the infection, where they expressed high levels of inflammation-induced cell-activation blocker PD-1, and became incapable of migrating back to the tumor site. At the same time, co-infection virus-specific CD8+ T cells remained functional while the infection was cleared. It was also unexpectedly found that PD-1 blockade immunotherapy reversed this effect. Here, we proceed to ground the experimental observations in a mechanistic immunobiochemical model that incorporates T cell pathways that control PD-1 expression. A core component of our model is a kinetic motif, which we call a PD-1 Double Incoherent Feed-Forward Loop (DIFFL), and which reflects known interactions between IRF4, Blimp-1, and Bcl-6. The different activity levels of the PD-1 DIFFL components, as a function of the cognate antigen levels and the given inflammation context, manifest themselves in phenotypically distinct outcomes. Collectively, the model allowed us to put forward a few working hypotheses as follows: (i) the melanoma-specific CD8+ T cells re-circulating with the blood flow enter the lung where they express high levels of inflammation-induced cell-activation blocker PD-1 in the presence of infection; (ii) when PD-1 receptors interact with abundant PD-L1, constitutively expressed in the lung, T cells loose motility; (iii) at the same time, virus-specific cells adapt to strong stimulation by their cognate antigen by lowering the transiently-elevated expression of PD-1, remaining functional and mobile in the inflamed lung, while the infection is cleared. The role that T cell receptor (TCR) activation and feedback loops play in the underlying processes are also highlighted and discussed. We hope that the results reported in our study could potentially contribute to the advancement of immunological approaches to cancer treatment and, as well, to a better understanding of a broader complexity of fundamental interactions between pathogens and tumors.
Keywords: PD-1/PD-L1; incoherent feedforward loop; influenza; mathematical modeling; melanoma.
Figures







Similar articles
-
Tumor immune profiling predicts response to anti-PD-1 therapy in human melanoma.J Clin Invest. 2016 Sep 1;126(9):3447-52. doi: 10.1172/JCI87324. Epub 2016 Aug 15. J Clin Invest. 2016. PMID: 27525433 Free PMC article.
-
Reactive glia promote development of CD103+ CD69+ CD8+ T-cells through programmed cell death-ligand 1 (PD-L1).Immun Inflamm Dis. 2018 Jun;6(2):332-344. doi: 10.1002/iid3.221. Epub 2018 Mar 30. Immun Inflamm Dis. 2018. PMID: 29602245 Free PMC article.
-
Non-oncogenic Acute Viral Infections Disrupt Anti-cancer Responses and Lead to Accelerated Cancer-Specific Host Death.Cell Rep. 2016 Oct 18;17(4):957-965. doi: 10.1016/j.celrep.2016.09.068. Cell Rep. 2016. PMID: 27760326 Free PMC article.
-
PD-L1 and immune escape: insights from melanoma and other lineage-unrelated malignancies.Hum Pathol. 2017 Aug;66:13-33. doi: 10.1016/j.humpath.2017.06.012. Epub 2017 Jul 8. Hum Pathol. 2017. PMID: 28694003 Review.
-
Cancer Cell-Intrinsic PD-1 and Implications in Combinatorial Immunotherapy.Front Immunol. 2018 Jul 30;9:1774. doi: 10.3389/fimmu.2018.01774. eCollection 2018. Front Immunol. 2018. PMID: 30105035 Free PMC article. Review.
Cited by
-
Immunological Paradigms, Mechanisms, and Models: Conceptual Understanding Is a Prerequisite to Effective Modeling.Front Immunol. 2019 Nov 5;10:2522. doi: 10.3389/fimmu.2019.02522. eCollection 2019. Front Immunol. 2019. PMID: 31749803 Free PMC article. Review.
-
Distributed Adaptive Search in T Cells: Lessons From Ants.Front Immunol. 2019 Jun 13;10:1357. doi: 10.3389/fimmu.2019.01357. eCollection 2019. Front Immunol. 2019. PMID: 31263465 Free PMC article. Review.
-
Delicate Balances in Cancer Chemotherapy: Modeling Immune Recruitment and Emergence of Systemic Drug Resistance.Front Immunol. 2020 Jun 30;11:1376. doi: 10.3389/fimmu.2020.01376. eCollection 2020. Front Immunol. 2020. PMID: 32695118 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous