

A Mitochondrial Progesterone Receptor Increases Cardiac Beta-Oxidation and Remodeling
- PMID: 30746505
- PMCID: PMC6364628
- DOI: 10.1210/js.2018-00219
A Mitochondrial Progesterone Receptor Increases Cardiac Beta-Oxidation and Remodeling
Abstract
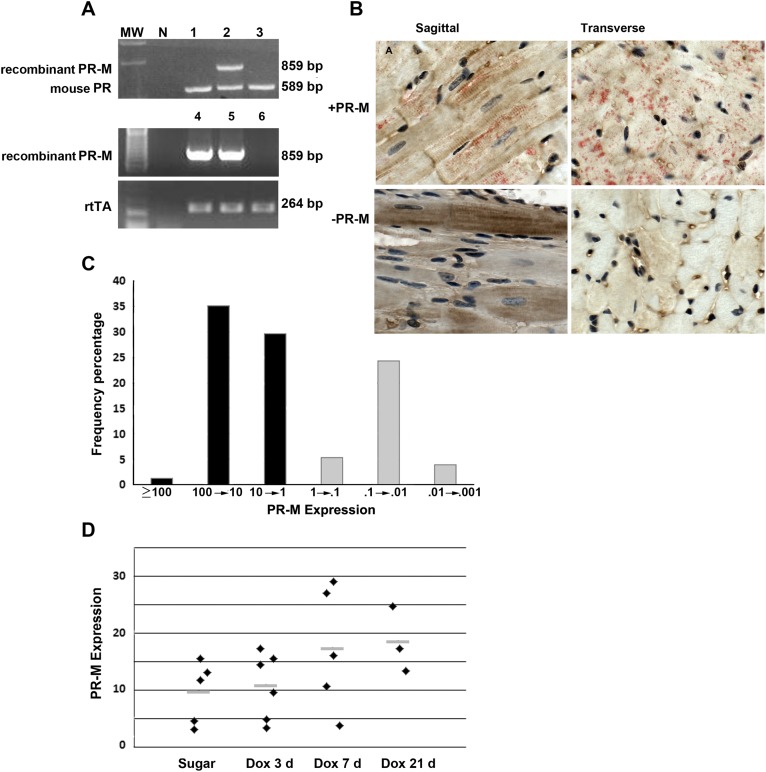
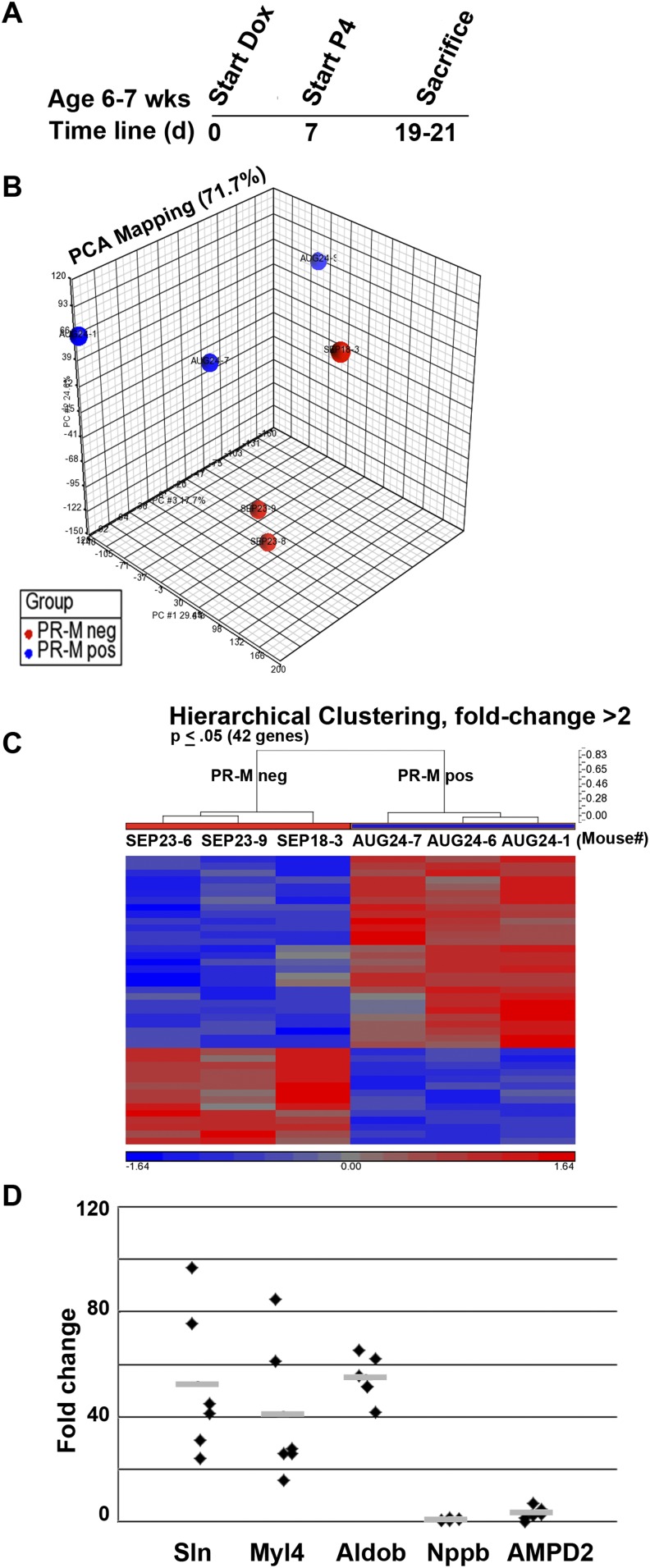
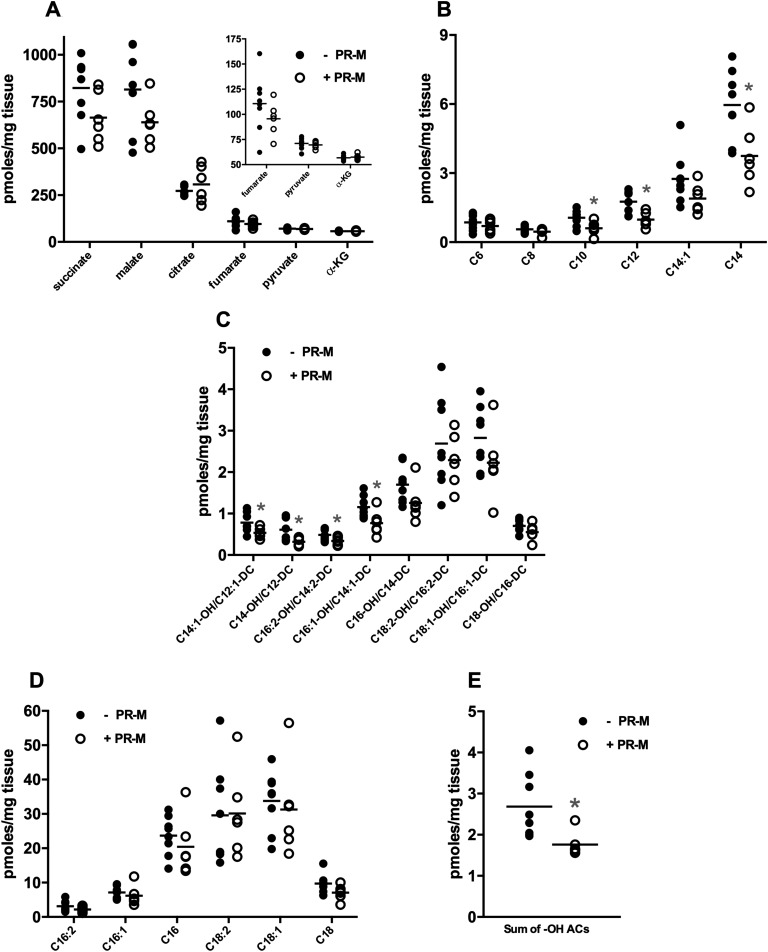
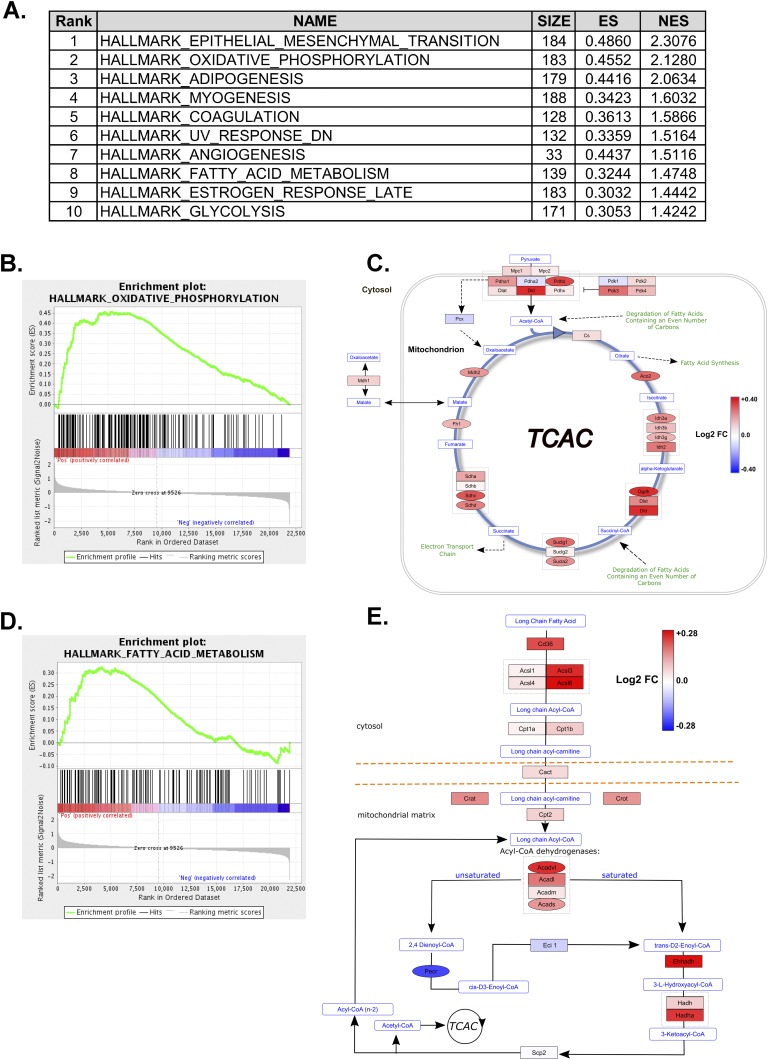
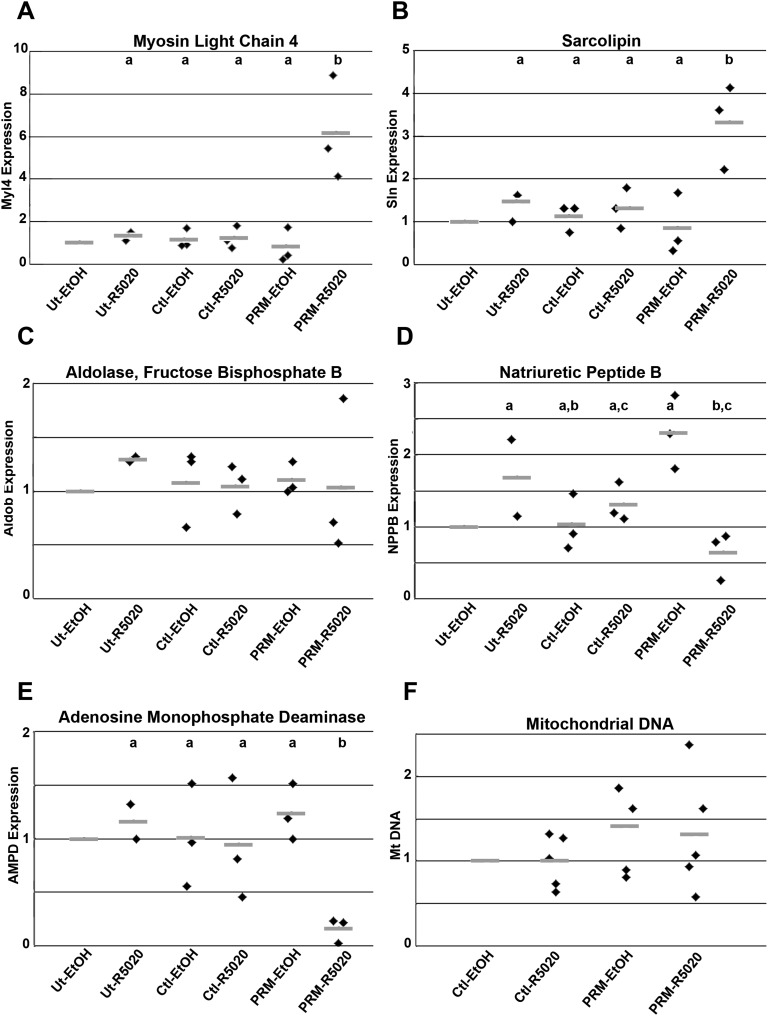
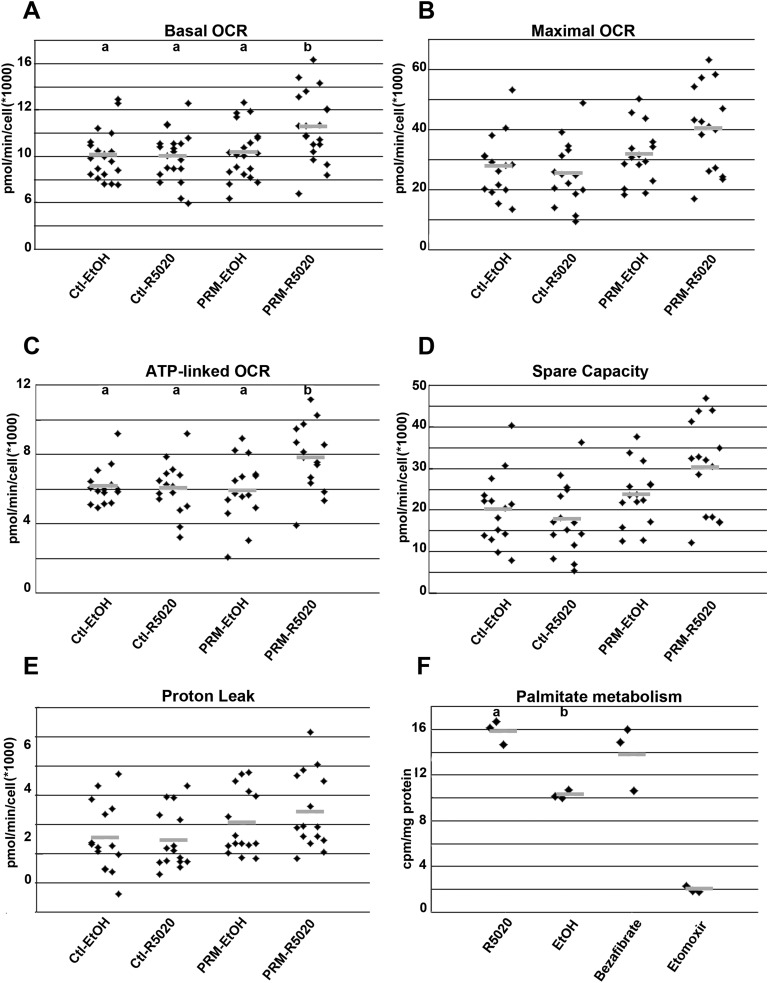
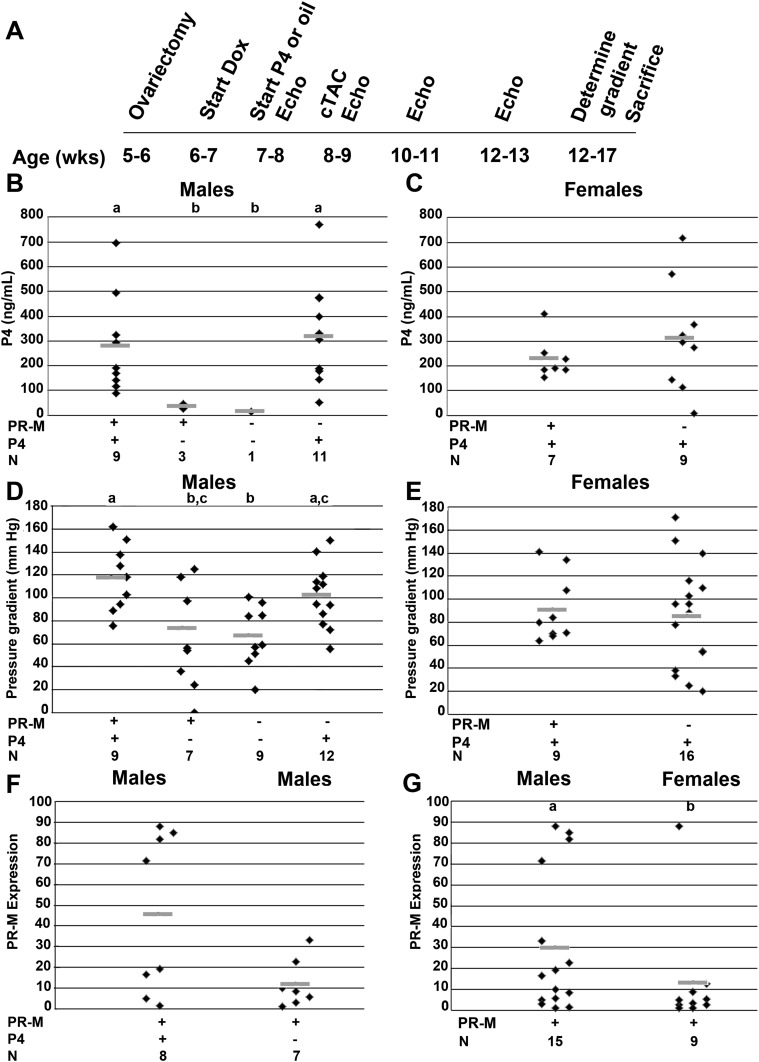
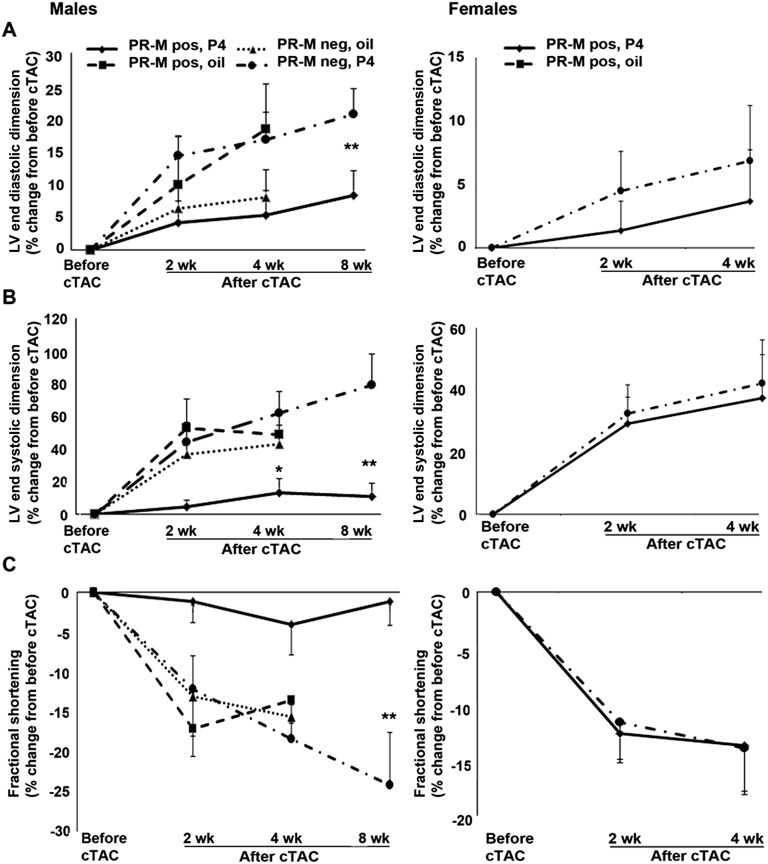
Progesterone is primarily a pregnancy-related hormone, produced in substantial quantities after ovulation and during gestation. Traditionally known to function via nuclear receptors for transcriptional regulation, there is also evidence of nonnuclear action. A previously identified mitochondrial progesterone receptor (PR-M) increases cellular respiration in cell models. In these studies, we demonstrated that expression of PR-M in rat H9c2 cardiomyocytes resulted in a ligand-dependent increase in oxidative cellular respiration and beta-oxidation. Cardiac expression in a TET-On transgenic mouse resulted in gene expression of myofibril proteins for remodeling and proteins involved in oxidative phosphorylation and fatty acid metabolism. In a model of increased afterload from constant transverse aortic constriction, mice expressing PR-M showed a ligand-dependent preservation of cardiac function. From these observations, we propose that PR-M is responsible for progesterone-induced increases in cellular energy production and cardiac remodeling to meet the physiological demands of pregnancy.
Keywords: beta-oxidation; mitochondria; pregnancy; progesterone; respiratory chain.
Figures
References
-
- Ouzounian JG, Elkayam U. Physiologic changes during normal pregnancy and delivery. Cardiol Clin. 2012;30(3):317–329. - PubMed
-
- Foryst-Ludwig A, Kintscher U. Sex differences in exercise-induced cardiac hypertrophy. Pflugers Arch. 2013;465(5):731–737. - PubMed
-
- Melchiorre K, Sutherland GR, Liberati M, Thilaganathan B. Preeclampsia is associated with persistent postpartum cardiovascular impairment. Hypertension. 2011;58(4):709–715. - PubMed
-
- Demakis JG, Rahimtoola SH, Sutton GC, Meadows WR, Szanto PB, Tobin JR, Gunnar RM. Natural course of peripartum cardiomyopathy. Circulation. 1971;44(6):1053–1061. - PubMed
-
- Lopaschuk GD, Kelly DP. Signalling in cardiac metabolism. Cardiovasc Res. 2008;79(2):205–207. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials