

The trafficking protein JFC1 regulates Rac1-GTP localization at the uropod controlling neutrophil chemotaxis and in vivo migration
- PMID: 30748033
- PMCID: PMC6543538
- DOI: 10.1002/JLB.1VMA0818-320R
The trafficking protein JFC1 regulates Rac1-GTP localization at the uropod controlling neutrophil chemotaxis and in vivo migration
Abstract
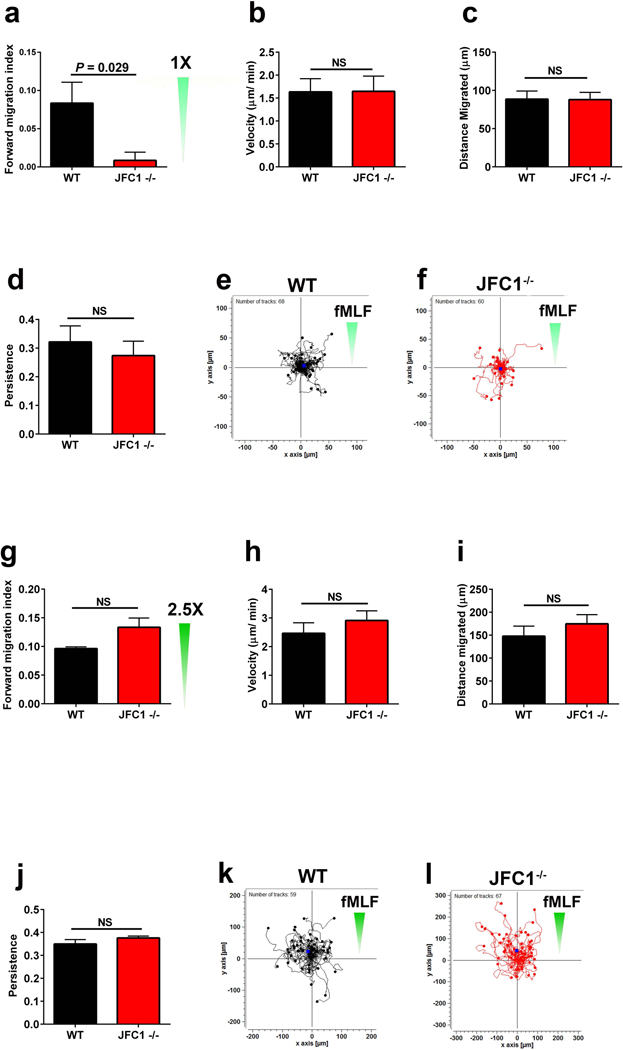
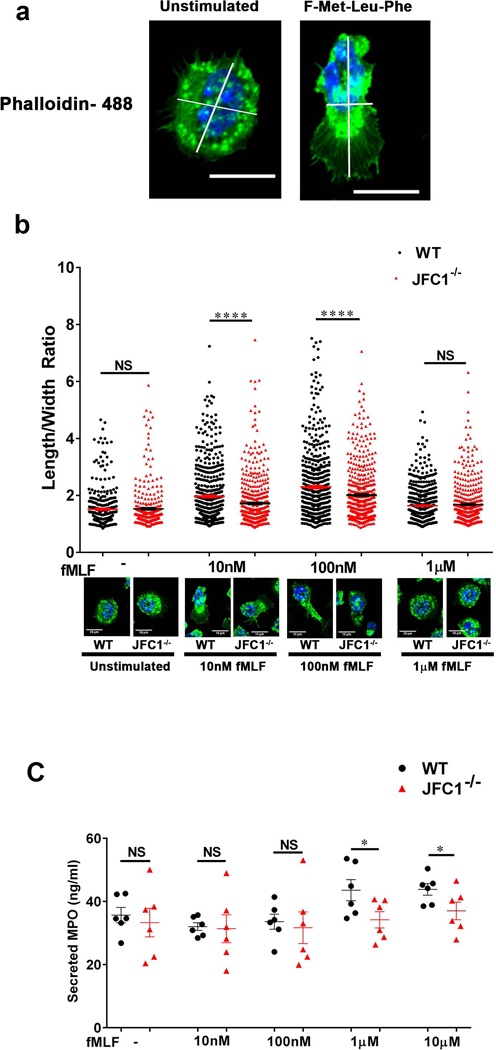
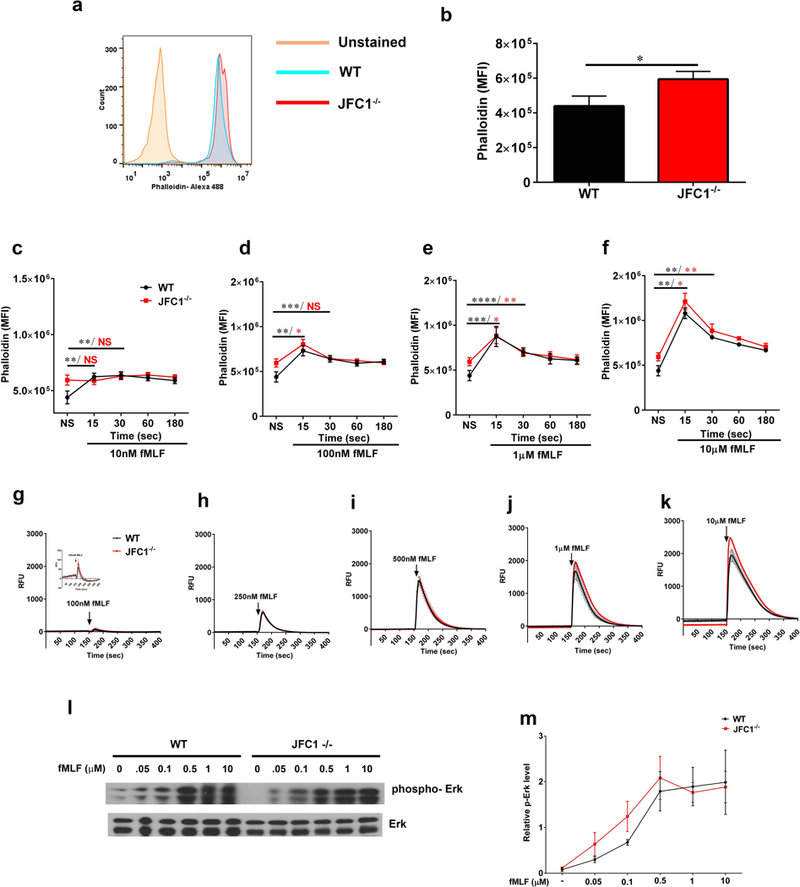
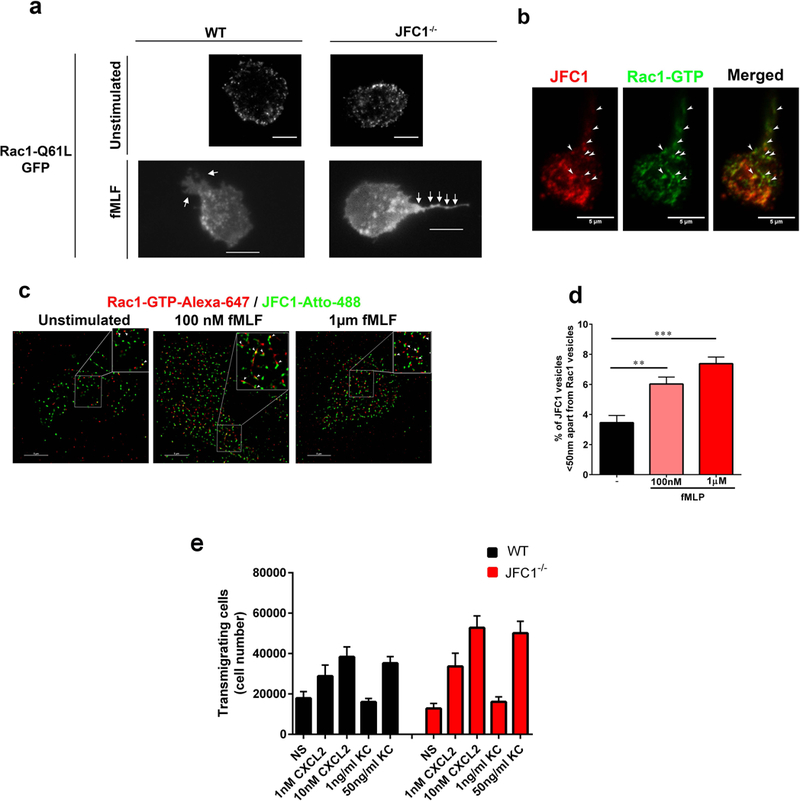
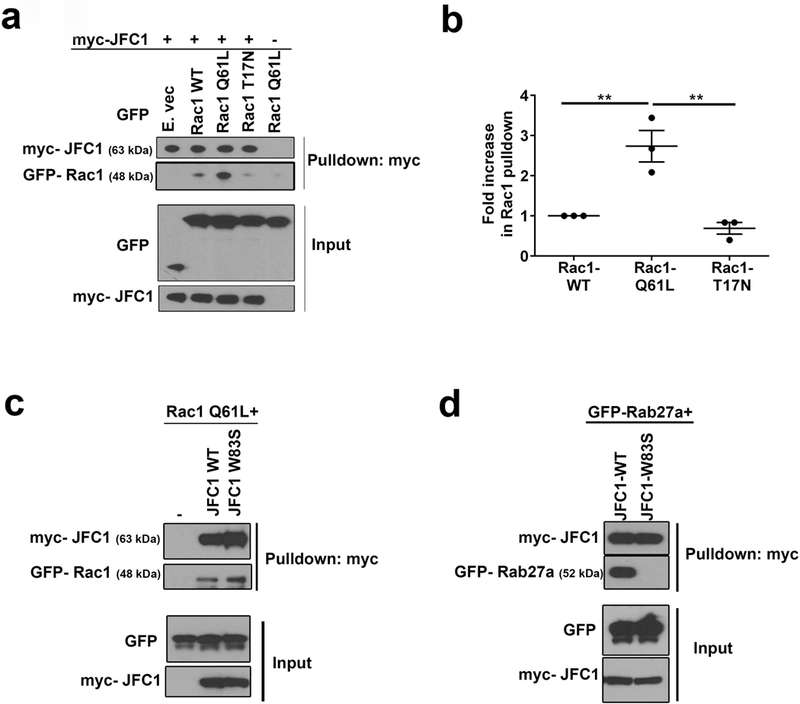
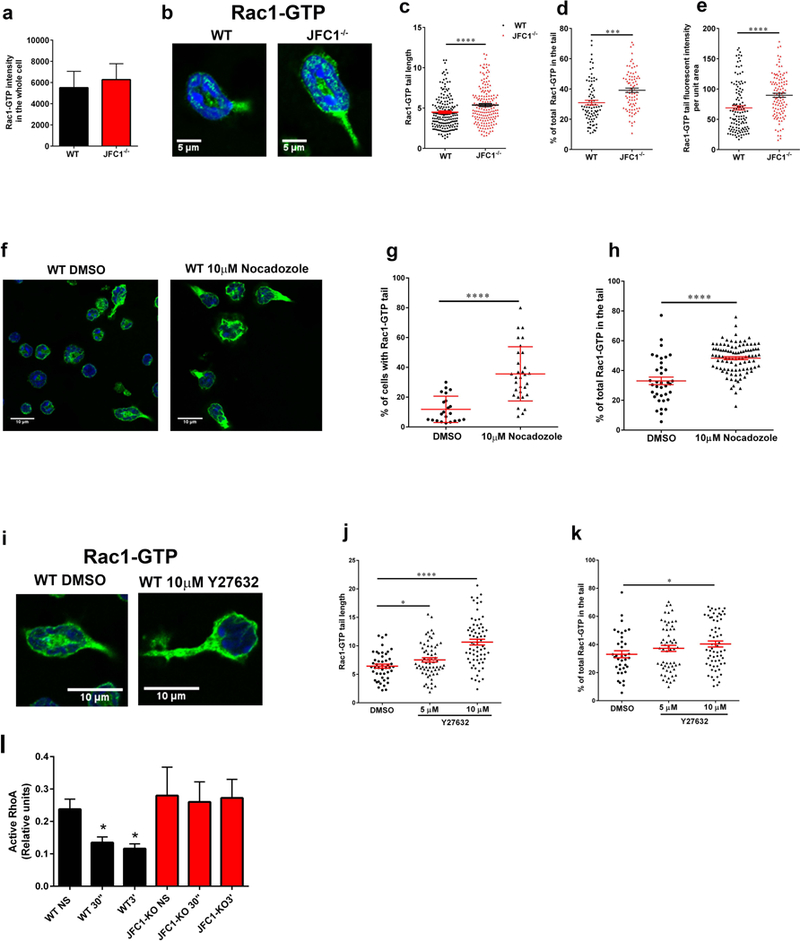
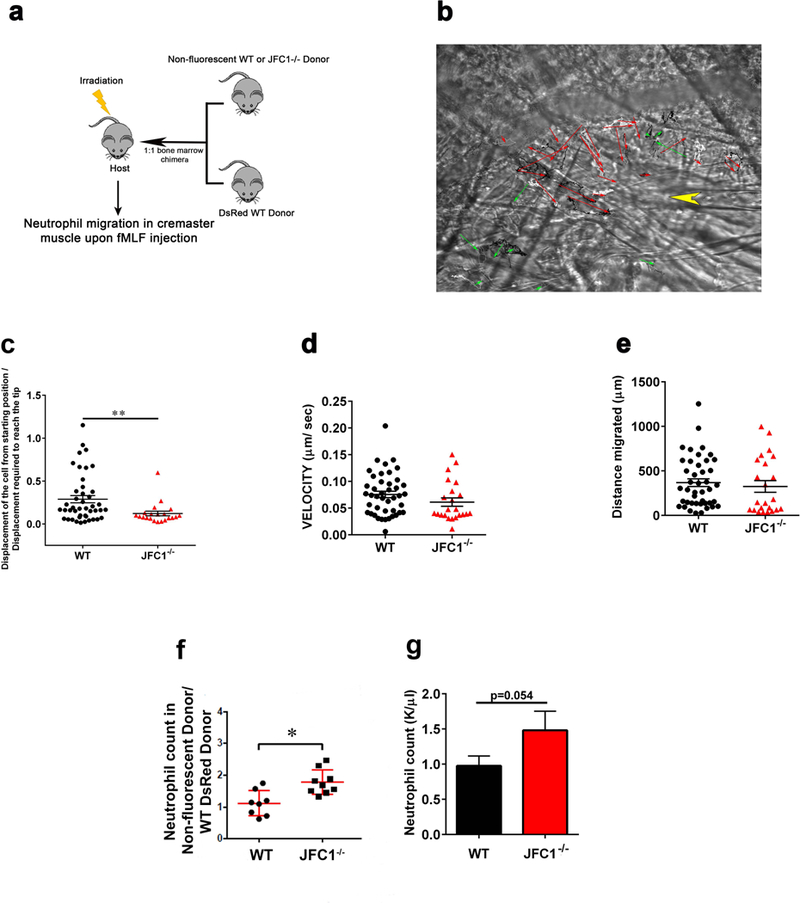
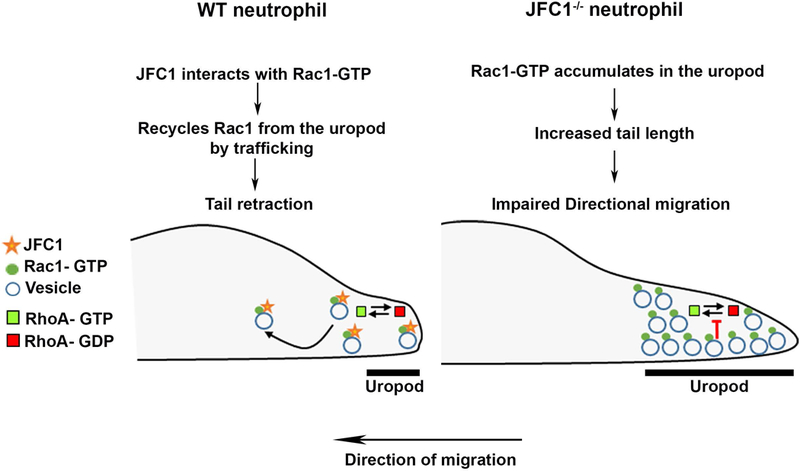
Neutrophil chemotaxis is essential in responses to infection and underlies inflammation. In neutrophils, the small GTPase Rac1 has discrete functions at both the leading edge and in the retraction of the trailing structure at the cell's rear (uropod), but how Rac1 is regulated at the uropod is unknown. Here, we identified a mechanism mediated by the trafficking protein synaptotagmin-like 1 (SYTL1 or JFC1) that controls Rac1-GTP recycling from the uropod and promotes directional migration of neutrophils. JFC1-null neutrophils displayed defective polarization and impaired directional migration to N-formyl-methionine-leucyl-phenylalanine in vitro, but chemoattractant-induced actin remodeling, calcium signaling and Erk activation were normal in these cells. Defective chemotaxis was not explained by impaired azurophilic granule exocytosis associated with JFC1 deficiency. Mechanistically, we show that active Rac1 localizes at dynamic vesicles where endogenous JFC1 colocalizes with Rac1-GTP. Super-resolution microscopy (STORM) analysis shows adjacent distribution of JFC1 and Rac1-GTP, which increases upon activation. JFC1 interacts with Rac1-GTP in a Rab27a-independent manner to regulate Rac1-GTP trafficking. JFC1-null cells exhibited Rac1-GTP accumulation at the uropod and increased tail length, and Rac1-GTP uropod accumulation was recapitulated by inhibition of ROCK or by interference with microtubule remodeling. In vivo, neutrophil dynamic studies in mixed bone marrow chimeric mice show that JFC1-/- neutrophils are unable to move directionally toward the source of the chemoattractant, supporting the notion that JFC1 deficiency results in defective neutrophil migration. Our results suggest that defective Rac1-GTP recycling from the uropod affects directionality and highlight JFC1-mediated Rac1 trafficking as a potential target to regulate chemotaxis in inflammation and immunity.
Keywords: JFC1; Rac1; chemotaxis; neutrophils; vesicular trafficking.
©2019 Society for Leukocyte Biology.
Conflict of interest statement
Figures
References
-
- Pertz O (2010) Spatio-temporal Rho GTPase signaling - where are we now? J Cell Sci 123, 1841–1850 - PubMed
-
- Wong CH, Heit B, and Kubes P (2010) Molecular regulators of leucocyte chemotaxis during inflammation. Cardiovascular research 86, 183–191 - PubMed
-
- Sun R, Iribarren P, Zhang N, Zhou Y, Gong W, Cho EH, Lockett S, Chertov O, Bednar F, Rogers TJ, Oppenheim JJ, and Wang JM (2004) Identification of neutrophil granule protein cathepsin G as a novel chemotactic agonist for the G protein-coupled formyl peptide receptor. J Immunol 173, 428–436 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
