

Type I interferon protects neurons from prions in in vivo models
- PMID: 30753318
- PMCID: PMC6439327
- DOI: 10.1093/brain/awz016
Type I interferon protects neurons from prions in in vivo models
Abstract
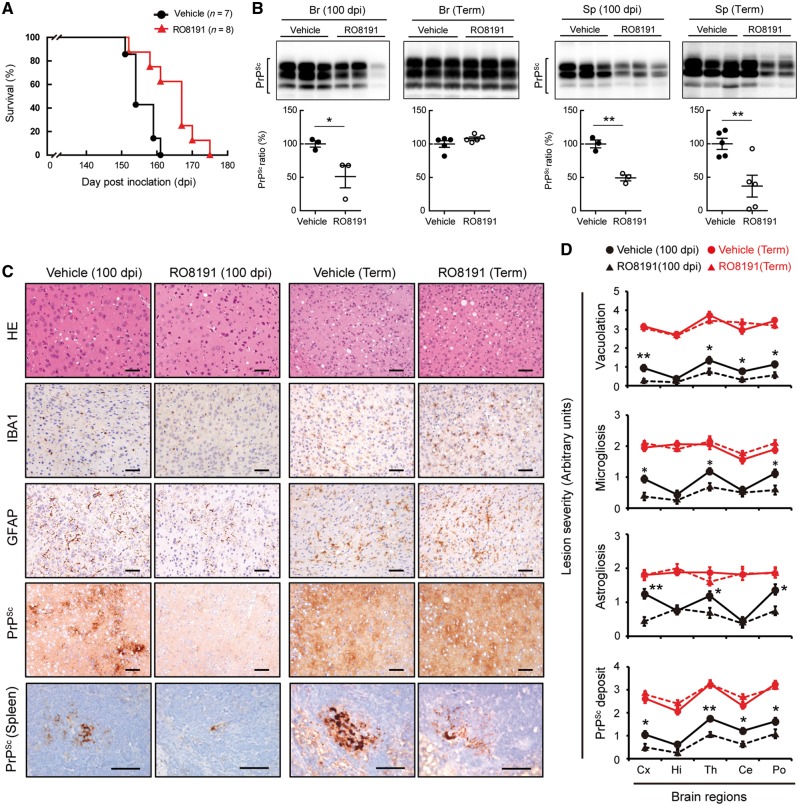
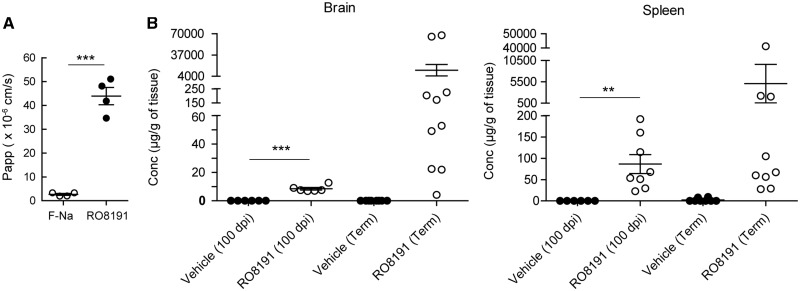
Infectious prions comprising abnormal prion protein, which is produced by structural conversion of normal prion protein, are responsible for transmissible spongiform encephalopathies including Creutzfeldt-Jakob disease in humans. Prions are infectious agents that do not possess a genome and the pathogenic protein was not thought to evoke any immune response. Although we previously reported that interferon regulatory factor 3 (IRF3) was likely to be involved in the pathogenesis of prion diseases, suggesting the protective role of host innate immune responses mediated by IRF3 signalling, this remained to be clarified. Here, we investigated the reciprocal interactions of type I interferon evoked by IRF3 activation and prion infection and found that infecting prions cause the suppression of endogenous interferon expression. Conversely, treatment with recombinant interferons in an ex vivo model was able to inhibit prion infection. In addition, cells and mice deficient in type I interferon receptor (subunit interferon alpha/beta receptor 1), exhibited higher susceptibility to 22L-prion infection. Moreover, in in vivo and ex vivo prion-infected models, treatment with RO8191, a selective type I interferon receptor agonist, inhibited prion invasion and prolonged the survival period of infected mice. Taken together, these data indicated that the interferon signalling interferes with prion propagation and some interferon-stimulated genes might play protective roles in the brain. These findings may allow for the development of new strategies to combat fatal diseases.
Keywords: innate immune system; prion infection; type I interferon (I-IFN).
© The Author(s) (2019). Published by Oxford University Press on behalf of the Guarantors of Brain.
Figures





Similar articles
-
[Protective Role of the Host Innate Immune System in Prion Pathogenesis].Yakugaku Zasshi. 2019;139(7):993-998. doi: 10.1248/yakushi.18-00165-2. Yakugaku Zasshi. 2019. PMID: 31257258 Review. Japanese.
-
THαβ Immunological Pathway as Protective Immune Response against Prion Diseases: An Insight for Prion Infection Therapy.Viruses. 2022 Feb 17;14(2):408. doi: 10.3390/v14020408. Viruses. 2022. PMID: 35216001 Free PMC article. Review.
-
Protective role of interferon regulatory factor 3-mediated signaling against prion infection.J Virol. 2012 May;86(9):4947-55. doi: 10.1128/JVI.06326-11. Epub 2012 Feb 29. J Virol. 2012. PMID: 22379081 Free PMC article.
-
Prion Protein Devoid of the Octapeptide Repeat Region Delays Bovine Spongiform Encephalopathy Pathogenesis in Mice.J Virol. 2017 Dec 14;92(1):e01368-17. doi: 10.1128/JVI.01368-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29046443 Free PMC article.
-
Protective role of MyD88-independent innate immune responses against prion infection.Prion. 2012 Nov-Dec;6(5):443-6. doi: 10.4161/pri.22579. Epub 2012 Oct 23. Prion. 2012. PMID: 23093799 Free PMC article.
Cited by
-
1-L Transcription in Prion Diseases.Int J Mol Sci. 2024 Sep 15;25(18):9961. doi: 10.3390/ijms25189961. Int J Mol Sci. 2024. PMID: 39337449 Free PMC article.
-
Reduced Expression of Prion Protein With Increased Interferon-β Fail to Limit Creutzfeldt-Jakob Disease Agent Replication in Differentiating Neuronal Cells.Front Physiol. 2022 Feb 18;13:837662. doi: 10.3389/fphys.2022.837662. eCollection 2022. Front Physiol. 2022. PMID: 35250638 Free PMC article.
-
Liquid-liquid phase separation of full-length prion protein initiates conformational conversion in vitro.J Biol Chem. 2021 Jan-Jun;296:100367. doi: 10.1016/j.jbc.2021.100367. Epub 2021 Feb 2. J Biol Chem. 2021. PMID: 33545172 Free PMC article.
-
Prion potentiation after life-long dormancy in mice devoid of PrP.Brain Commun. 2021 Apr 28;3(2):fcab092. doi: 10.1093/braincomms/fcab092. eCollection 2021. Brain Commun. 2021. PMID: 33997785 Free PMC article.
-
Type-I Interferons in Alzheimer's Disease and Other Tauopathies.Front Cell Neurosci. 2022 Jul 15;16:949340. doi: 10.3389/fncel.2022.949340. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35910253 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
