

ADAM10 sheddase activation is controlled by cell membrane asymmetry
- PMID: 30753537
- PMCID: PMC6927242
- DOI: 10.1093/jmcb/mjz008
ADAM10 sheddase activation is controlled by cell membrane asymmetry
Abstract
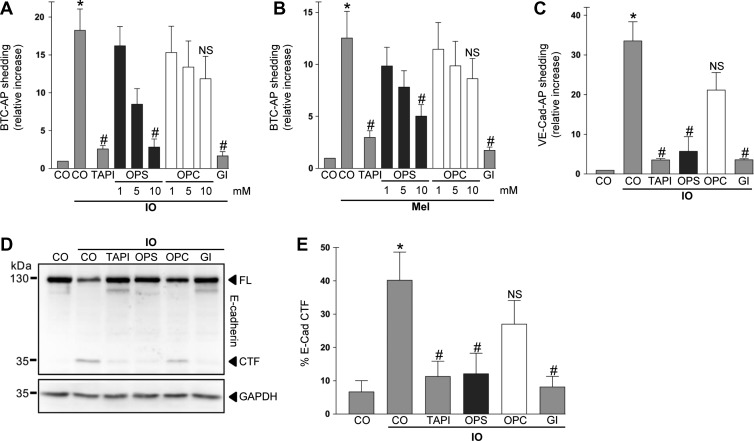
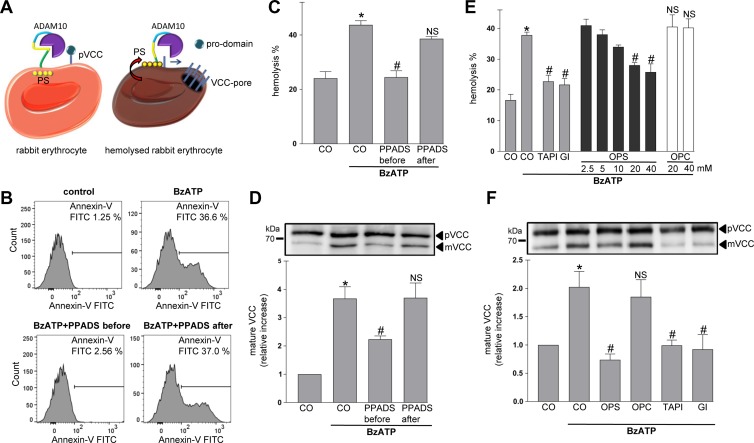
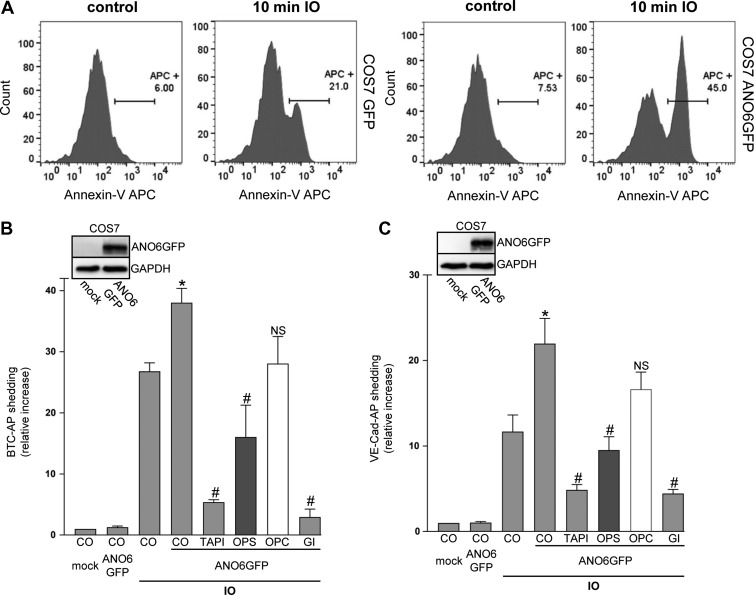
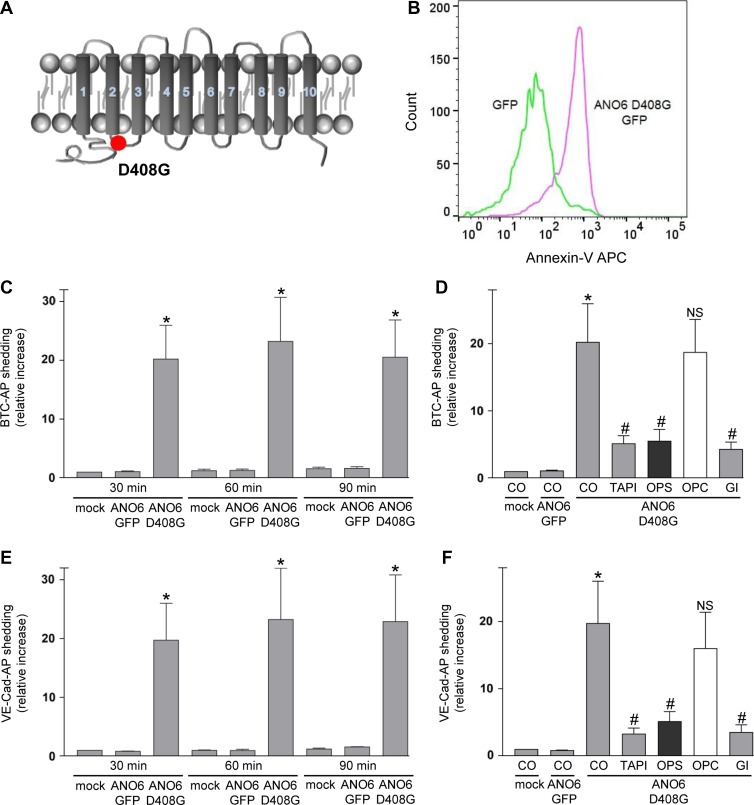
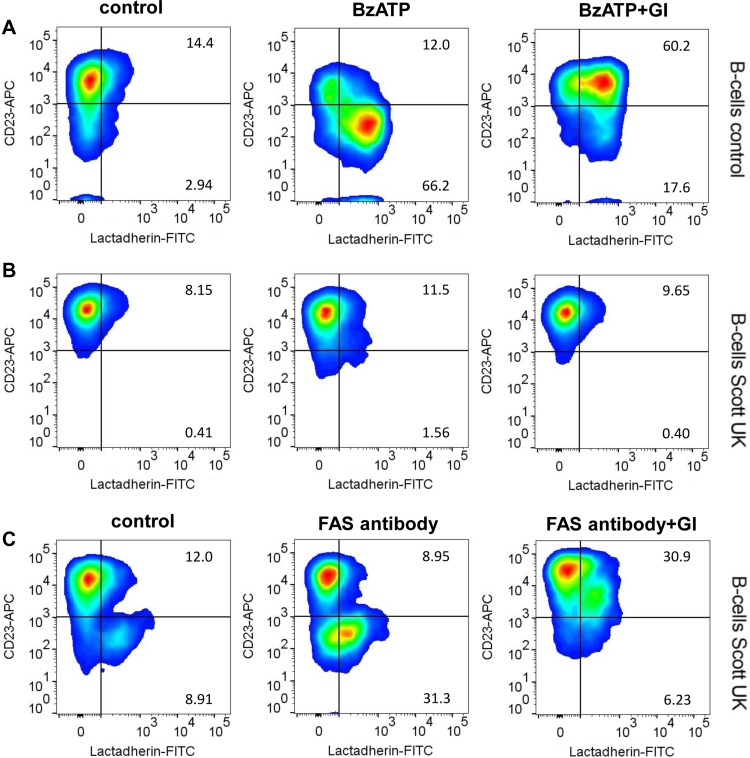
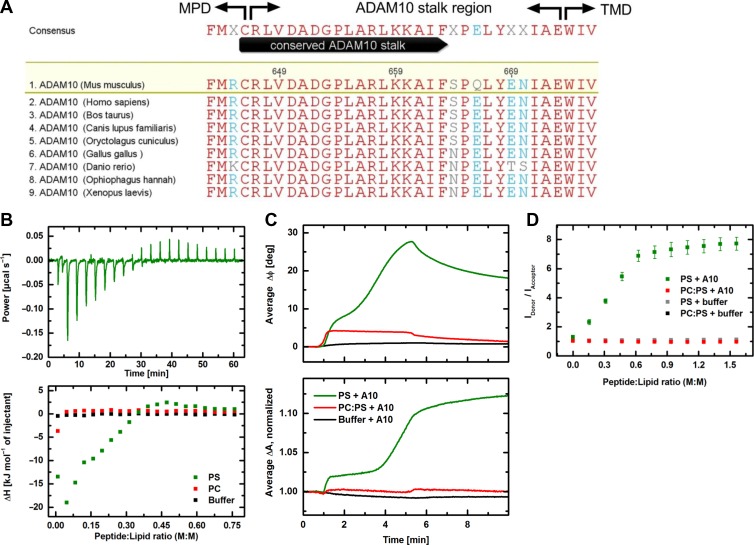
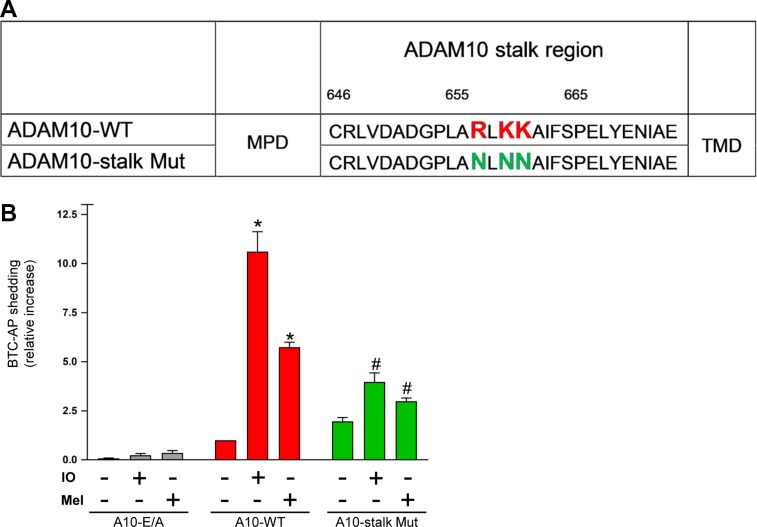
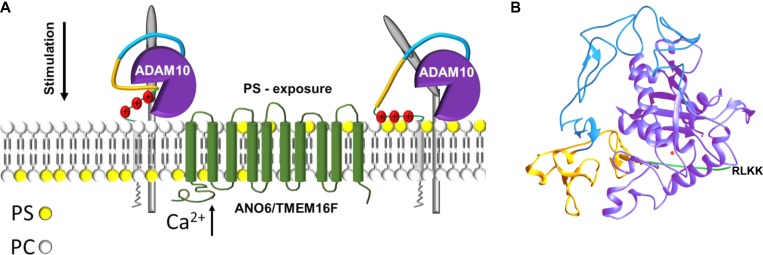
Dysregulation of the disintegrin-metalloproteinase ADAM10 may contribute to the development of diseases including tumorigenesis and Alzheimer's disease. The mechanisms underlying ADAM10 sheddase activation are incompletely understood. Here, we show that transient exposure of the negatively charged phospholipid phosphatidylserine (PS) is necessarily required. The soluble PS headgroup was found to act as competitive inhibitor of substrate cleavage. Overexpression of the Ca2+-dependent phospholipid scramblase Anoctamin-6 (ANO6) led to increased PS externalization and substrate release. Transfection with a constitutively active form of ANO6 resulted in maximum sheddase activity in the absence of any stimulus. Calcium-dependent ADAM10 activation could not be induced in lymphocytes of patients with Scott syndrome harbouring a missense mutation in ANO6. A putative PS-binding motif was identified in the conserved stalk region. Replacement of this motif resulted in strong reduction of sheddase activity. In conjunction with the recently described 3D structure of the ADAM10 extracellular domain, a model is advanced to explain how surface-exposed PS triggers ADAM10 sheddase function.
Keywords: ADAM10; Anoctamin-6; activation; cell membrane asymmetry; phosphatidylserine; shedding.
© The Author(s) (2019). Published by Oxford University Press on behalf of Journal of Molecular Cell Biology, IBCB, SIBS, CAS.
Figures
References
-
- Andra J., Bohling A., Gronewold T.M., et al. (2008). Surface acoustic wave biosensor as a tool to study the interaction of antimicrobial peptides with phospholipid and lipopolysaccharide model membranes. Langmuir 24, 9148–9153. - PubMed
-
- Bevers E.M., and Williamson P.L. (2016). Getting to the outer leaflet: physiology of phosphatidylserine exposure at the plasma membrane. Physiol. Rev. 96, 605–645. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
