

Loss of RNA-binding protein GRSF1 activates mTOR to elicit a proinflammatory transcriptional program
- PMID: 30753671
- PMCID: PMC6412117
- DOI: 10.1093/nar/gkz082
Loss of RNA-binding protein GRSF1 activates mTOR to elicit a proinflammatory transcriptional program
Abstract
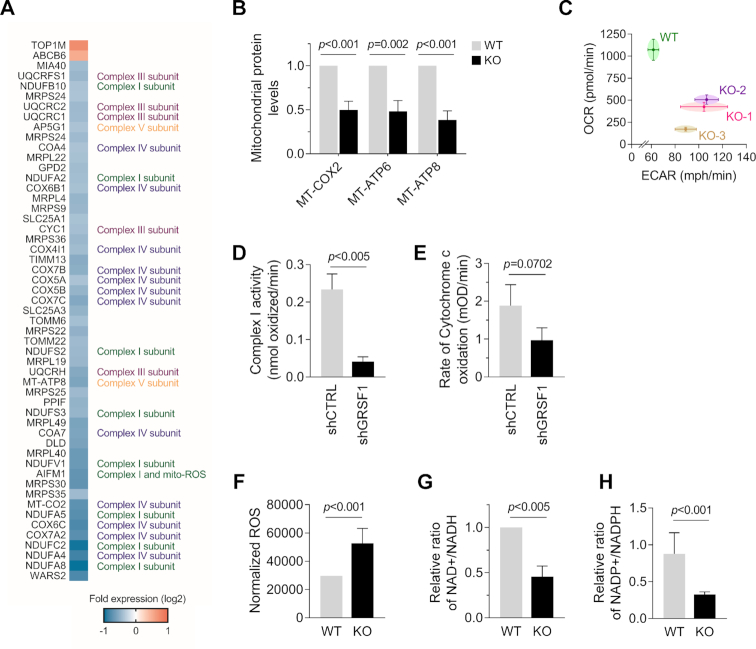
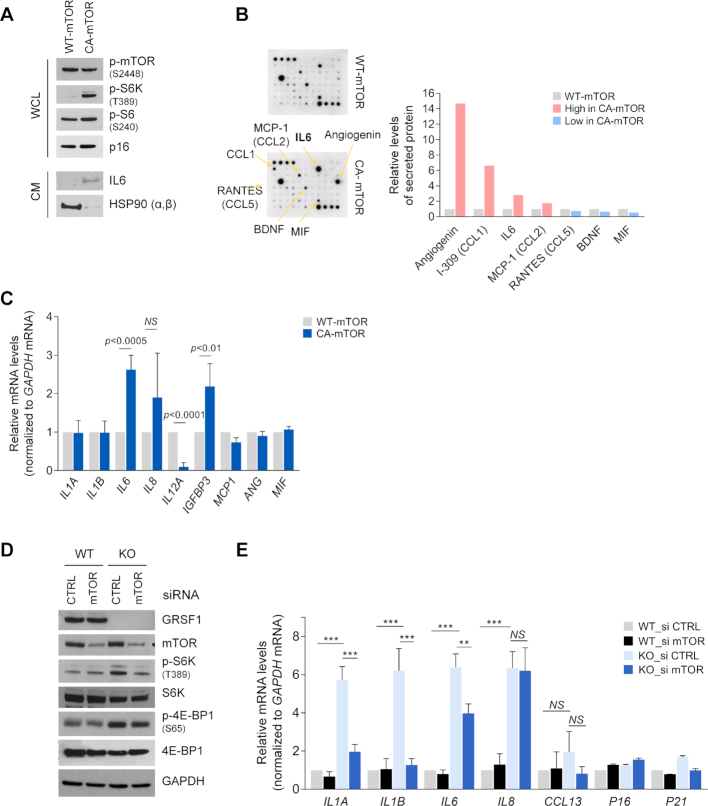
The RNA-binding protein GRSF1 (G-rich RNA sequence-binding factor 1) critically maintains mitochondrial homeostasis. Accordingly, loss of GRSF1 impaired mitochondrial respiration and increased the levels of reactive oxygen species (ROS), triggering DNA damage, growth suppression, and a senescent phenotype characterized by elevated production and secretion of interleukin (IL)6. Here, we characterize the pathways that govern IL6 production in response to mitochondrial dysfunction in GRSF1-depleted cells. We report that loss of GRSF1 broadly altered protein expression programs, impairing the function of respiratory complexes I and IV. The rise in oxidative stress led to increased DNA damage and activation of mTOR, which in turn activated NF-κB to induce IL6 gene transcription and orchestrate a pro-inflammatory program. Collectively, our results indicate that GRSF1 helps preserve mitochondrial homeostasis, in turn preventing oxidative DNA damage and the activation of mTOR and NF-κB, and suppressing a transcriptional pro-inflammatory program leading to increased IL6 production.
Published by Oxford University Press on behalf of Nucleic Acids Research 2019.
Figures





References
-
- Dreyfuss G., Kim V.N., Kataoka N.. Messenger RNA binding proteins and the messages they carry. Nat. Rev. Mol. Cell Biol. 2002; 3:195–205. - PubMed
-
- Keene J.D. RNA regulons: coordination of posttranscriptional events. Nat. Rev. Genet. 2007; 8:533–543. - PubMed
-
- Moore M.J. From birth to death: the complex lives of eukaryotic mRNAs. Science. 2005; 309:1514–1518. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
