

Root volatiles in plant-plant interactions II: Root volatiles alter root chemistry and plant-herbivore interactions of neighbouring plants
- PMID: 30754075
- PMCID: PMC6849603
- DOI: 10.1111/pce.13534
Root volatiles in plant-plant interactions II: Root volatiles alter root chemistry and plant-herbivore interactions of neighbouring plants
Abstract
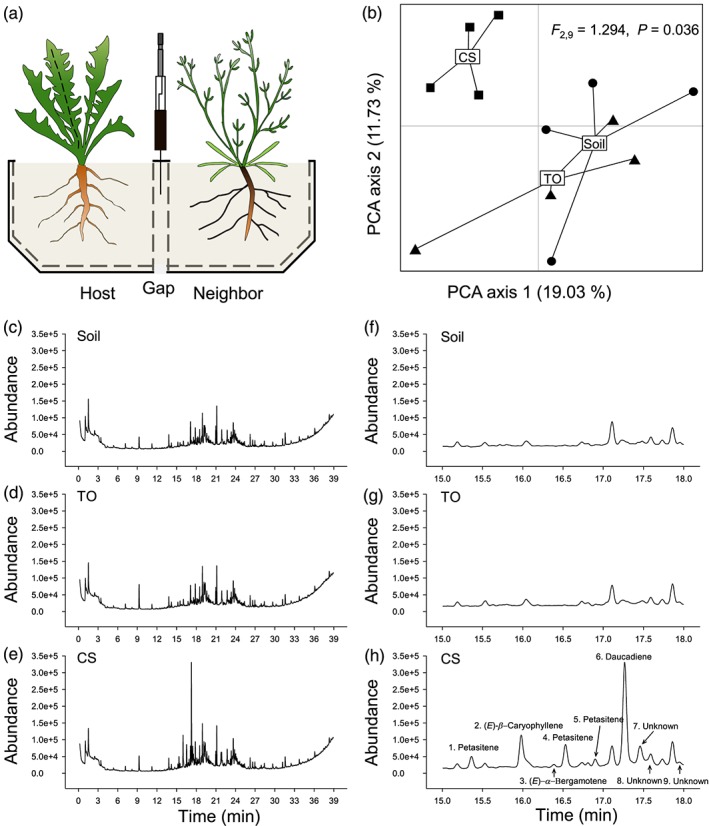
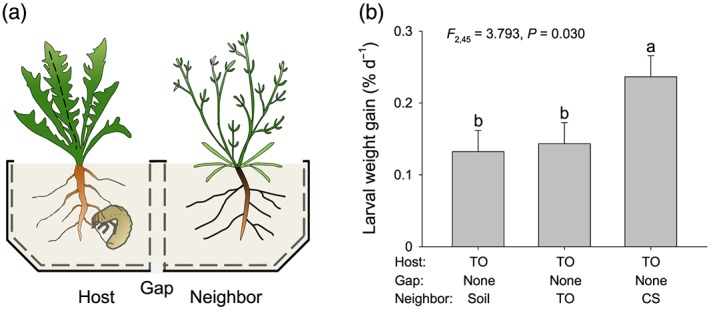
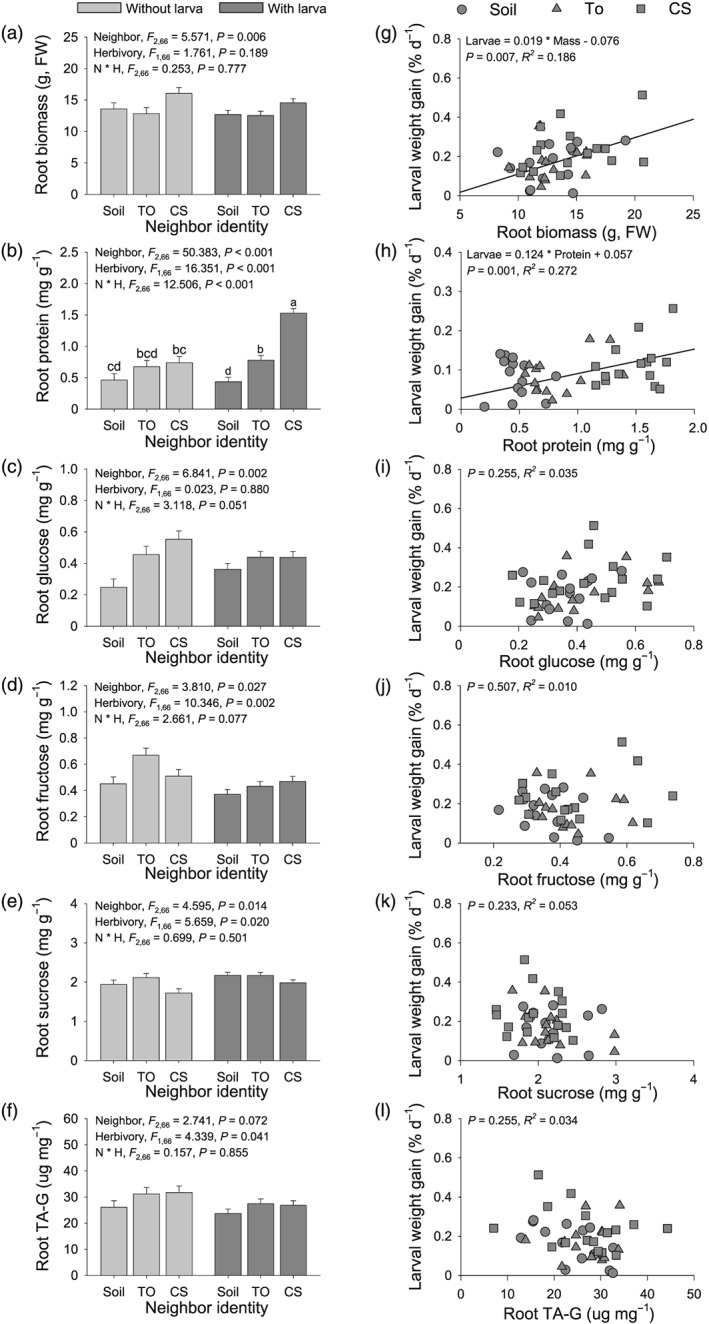
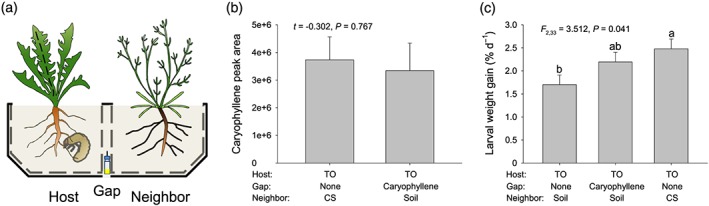
Volatile organic compounds (VOCs) emitted by plant roots can influence the germination and growth of neighbouring plants. However, little is known about the effects of root VOCs on plant-herbivore interactions of neighbouring plants. The spotted knapweed (Centaurea stoebe) constitutively releases high amounts of sesquiterpenes into the rhizosphere. Here, we examine the impact of C. stoebe root VOCs on the primary and secondary metabolites of sympatric Taraxacum officinale plants and the resulting plant-mediated effects on a generalist root herbivore, the white grub Melolontha melolontha. We show that exposure of T. officinale to C.stoebe root VOCs does not affect the accumulation of defensive secondary metabolites but modulates carbohydrate and total protein levels in T. officinale roots. Furthermore, VOC exposure increases M. melolontha growth on T. officinale plants. Exposure of T. officinale to a major C. stoebe root VOC, the sesquiterpene (E)-β-caryophyllene, partially mimics the effect of the full root VOC blend on M. melolontha growth. Thus, releasing root VOCs can modify plant-herbivore interactions of neighbouring plants. The release of VOCs to increase the susceptibility of other plants may be a form of plant offense.
Keywords: associational effects; belowground herbivory; neighbourhood effects; plant-herbivore interactions; plant-plant interactions; volatile priming.
© 2019 The Authors Plant, Cell & Environment Published by John Wiley & Sons Ltd.
Figures
References
-
- Arimura, G.‐I. , Ozawa, R. , Shimoda, T. , Nishioka, T. , Boland, W. , & Takabayashi, J. (2000). Herbivory‐induced volatiles elicit defence genes in lima bean leaves. Nature, 406, 512–515. - PubMed
-
- Barbosa, P. , Hines, J. , Kaplan, I. , Martinson, H. , Szczepaniec, A. , & Szendrei, Z. (2009). Associational resistance and associational susceptibility: Having right or wrong neighbors. Annual Review of Ecology, Evolution, and Systematics, 40, 1–20. 10.1146/annurev.ecolsys.110308.120242 - DOI
-
- Bates, D. , Mächler, M. , Bolker, B. , & Walker, S. (2015). Fitting linear mixed‐effects models using lme4. Journal of Statistical Software, 67, 1–48.
-
- Benjamini, Y. , & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B‐Statistical Methodology, 57, 289–300.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
