

PHF6 regulates hematopoietic stem and progenitor cells and its loss synergizes with expression of TLX3 to cause leukemia
- PMID: 30755422
- PMCID: PMC6695515
- DOI: 10.1182/blood-2018-07-860726
PHF6 regulates hematopoietic stem and progenitor cells and its loss synergizes with expression of TLX3 to cause leukemia
Abstract
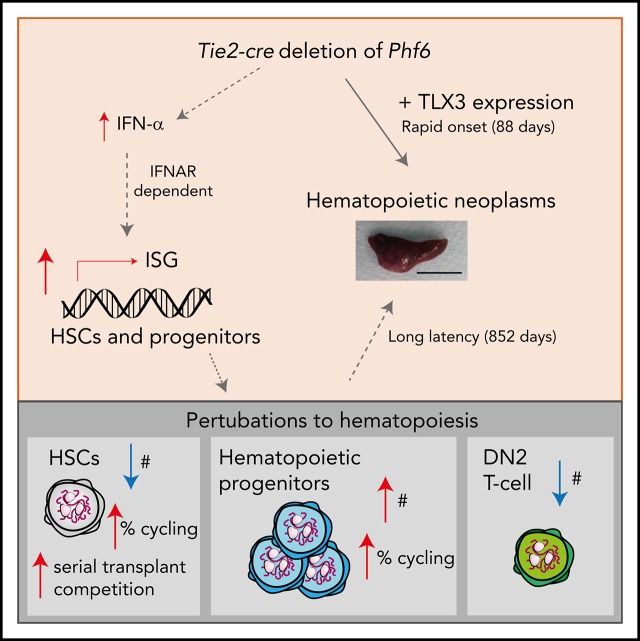
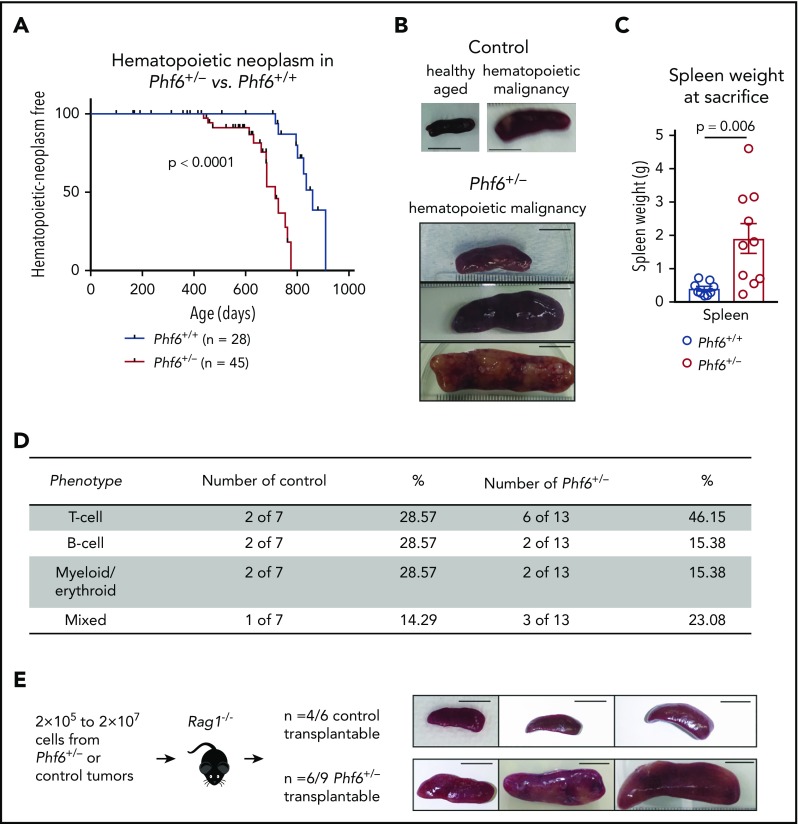
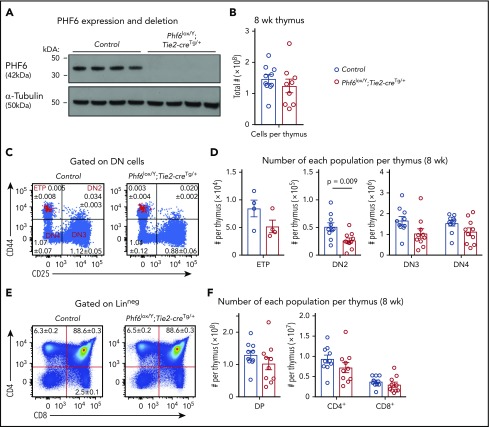
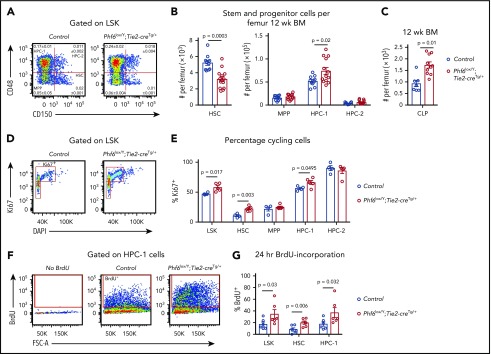
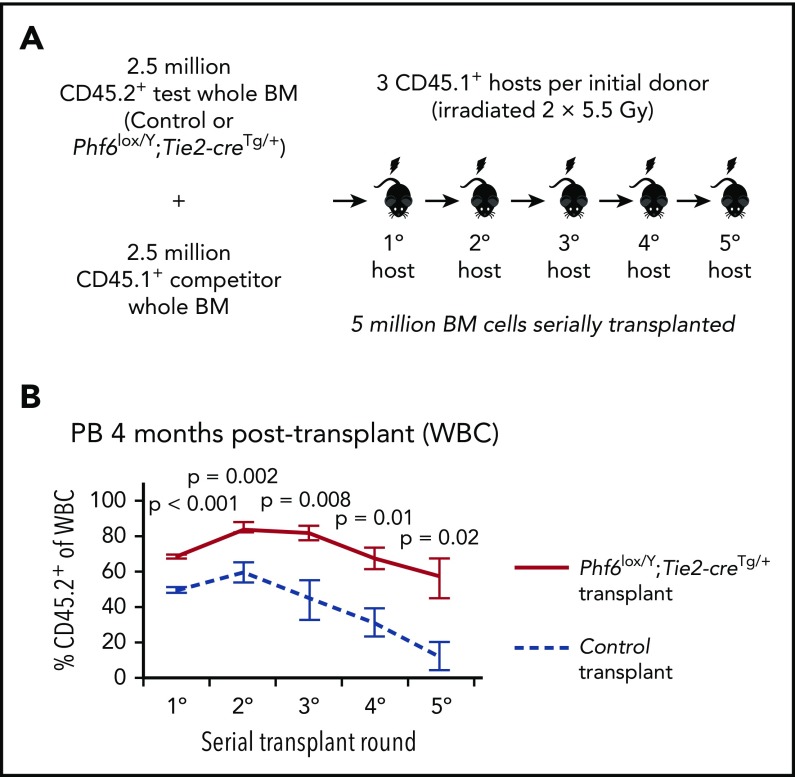
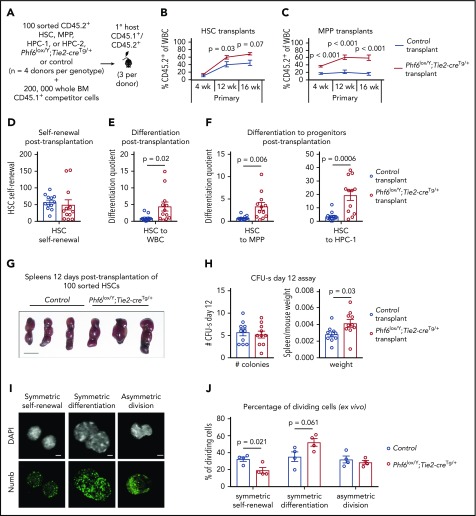
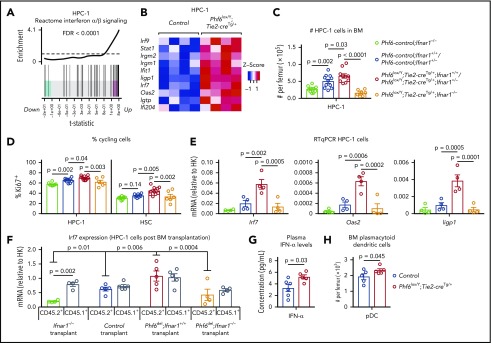
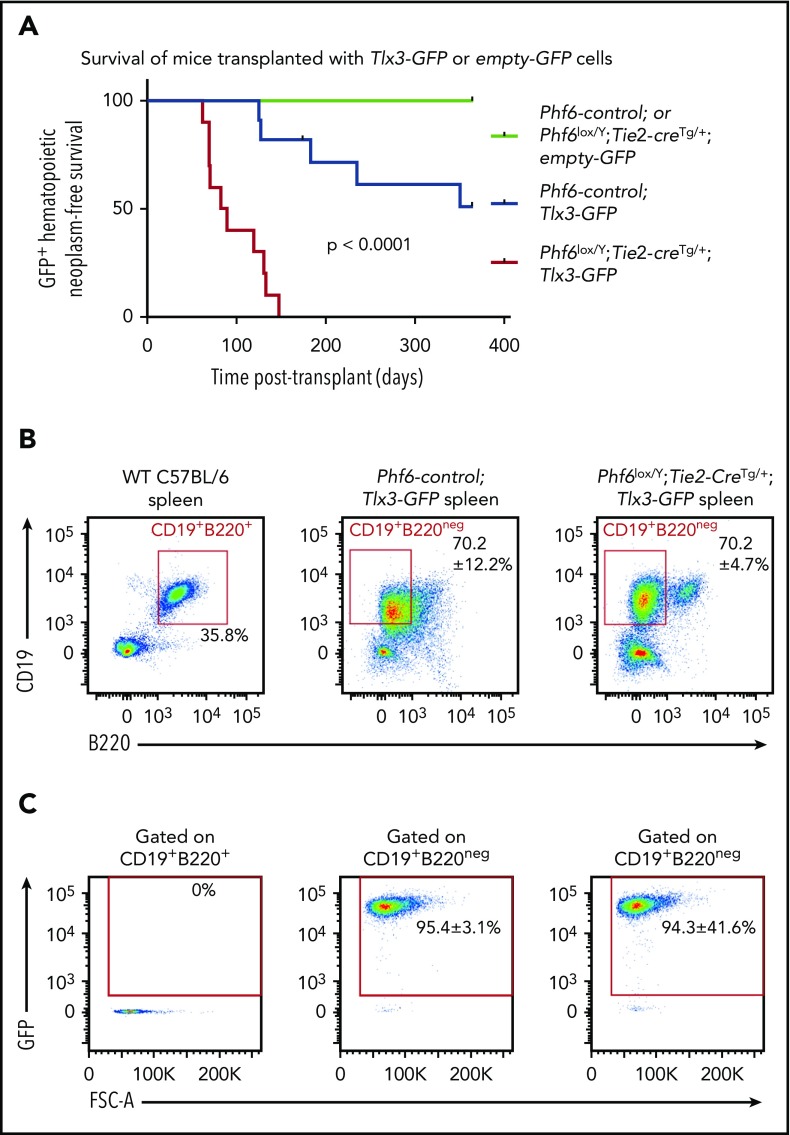
Somatically acquired mutations in PHF6 (plant homeodomain finger 6) frequently occur in hematopoietic malignancies and often coincide with ectopic expression of TLX3. However, there is no functional evidence to demonstrate whether these mutations contribute to tumorigenesis. Similarly, the role of PHF6 in hematopoiesis is unknown. We report here that Phf6 deletion in mice resulted in a reduced number of hematopoietic stem cells (HSCs), an increased number of hematopoietic progenitor cells, and an increased proportion of cycling stem and progenitor cells. Loss of PHF6 caused increased and sustained hematopoietic reconstitution in serial transplantation experiments. Interferon-stimulated gene expression was upregulated in the absence of PHF6 in hematopoietic stem and progenitor cells. The numbers of hematopoietic progenitor cells and cycling hematopoietic stem and progenitor cells were restored to normal by combined loss of PHF6 and the interferon α and β receptor subunit 1. Ectopic expression of TLX3 alone caused partially penetrant leukemia. TLX3 expression and loss of PHF6 combined caused fully penetrant early-onset leukemia. Our data suggest that PHF6 is a hematopoietic tumor suppressor and is important for fine-tuning hematopoietic stem and progenitor cell homeostasis.
© 2019 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Comment in
-
Context-specific tumor suppression by PHF6.Blood. 2019 Apr 18;133(16):1698-1700. doi: 10.1182/blood-2019-03-900829. Blood. 2019. PMID: 31000514 Free PMC article.
References
-
- Li X, Yao H, Chen Z, Wang Q, Zhao Y, Chen S. Somatic mutations of PHF6 in patients with chronic myeloid leukemia in blast crisis. Leuk Lymphoma. 2013;54(3):671-672. - PubMed
-
- Xiao W, Pastore F, Getta B, et al. PHF6 mutations defines a subgroup of mixed phenotype of acute leukemia with aberrant T-cell differentiation [abstract]. Blood. 2017;130(suppl 1). Abstract 1384.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
