

Long-Term Transcriptional Activity at Zero Growth of a Cosmopolitan Rare Biosphere Member
- PMID: 30755506
- PMCID: PMC6372793
- DOI: 10.1128/mBio.02189-18
Long-Term Transcriptional Activity at Zero Growth of a Cosmopolitan Rare Biosphere Member
Abstract
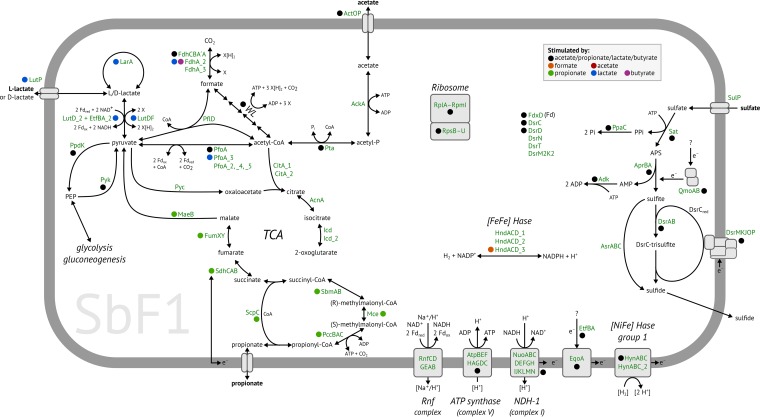
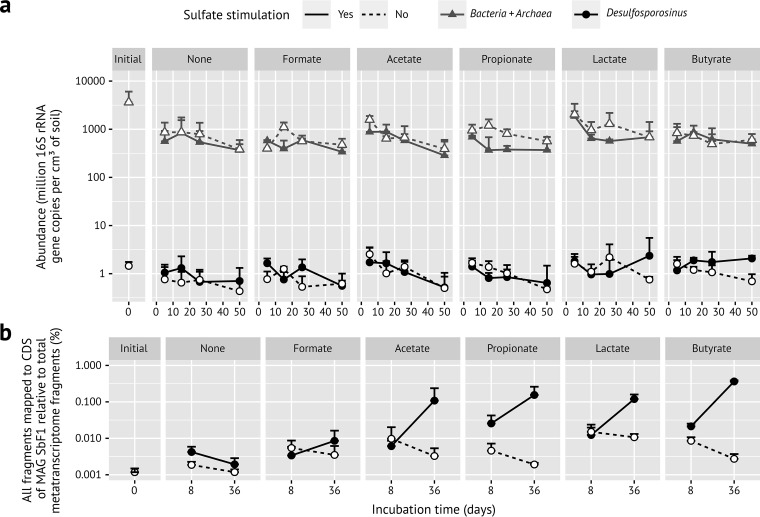
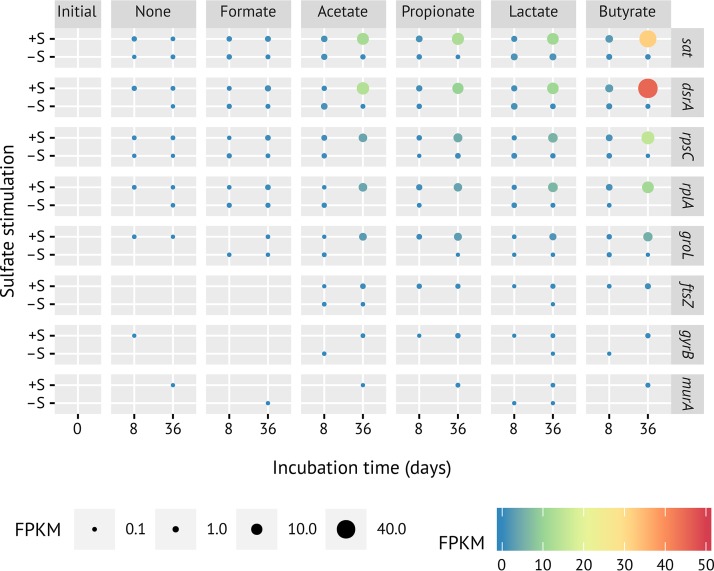
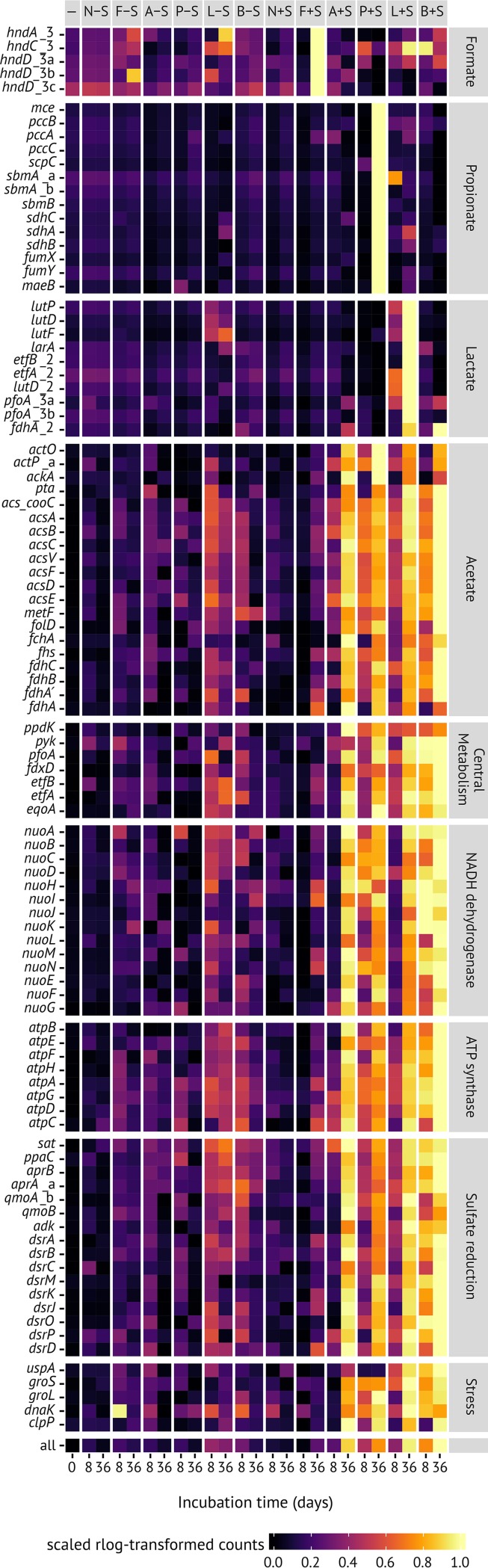
Microbial diversity in the environment is mainly concealed within the rare biosphere (all species with <0.1% relative abundance). While dormancy explains a low-abundance state very well, the mechanisms leading to rare but active microorganisms remain elusive. We used environmental systems biology to genomically and transcriptionally characterize "Candidatus Desulfosporosinus infrequens," a low-abundance sulfate-reducing microorganism cosmopolitan to freshwater wetlands, where it contributes to cryptic sulfur cycling. We obtained its near-complete genome by metagenomics of acidic peat soil. In addition, we analyzed anoxic peat soil incubated under in situ-like conditions for 50 days by Desulfosporosinus-targeted qPCR and metatranscriptomics. The Desulfosporosinus population stayed at a constant low abundance under all incubation conditions, averaging 1.2 × 106 16S rRNA gene copies per cm³ soil. In contrast, transcriptional activity of "Ca. Desulfosporosinus infrequens" increased at day 36 by 56- to 188-fold when minor amendments of acetate, propionate, lactate, or butyrate were provided with sulfate, compared to the no-substrate-control. Overall transcriptional activity was driven by expression of genes encoding ribosomal proteins, energy metabolism, and stress response but not by expression of genes encoding cell growth-associated processes. Since our results did not support growth of these highly active microorganisms in terms of biomass increase or cell division, they had to invest their sole energy for maintenance, most likely counterbalancing acidic pH conditions. This finding explains how a rare biosphere member can contribute to a biogeochemically relevant process while remaining in a zero-growth state over a period of 50 days.IMPORTANCE The microbial rare biosphere represents the largest pool of biodiversity on Earth and constitutes, in sum of all its members, a considerable part of a habitat's biomass. Dormancy or starvation is typically used to explain the persistence of low-abundance microorganisms in the environment. We show that a low-abundance microorganism can be highly transcriptionally active while remaining in a zero-growth state for at least 7 weeks. Our results provide evidence that this zero growth at a high cellular activity state is driven by maintenance requirements. We show that this is true for a microbial keystone species, in particular a cosmopolitan but permanently low-abundance sulfate-reducing microorganism in wetlands that is involved in counterbalancing greenhouse gas emissions. In summary, our results provide an important step forward in understanding time-resolved activities of rare biosphere members relevant for ecosystem functions.
Keywords: cryptic sulfur cycle; growth arrest; keystone species; maintenance; metatranscriptome; peatland.
Copyright © 2019 Hausmann et al.
Figures
Comment in
-
Life in the slow lane.Nat Rev Microbiol. 2019 May;17(5):266-267. doi: 10.1038/s41579-019-0176-7. Nat Rev Microbiol. 2019. PMID: 30833717 No abstract available.
References
-
- Jousset A, Bienhold C, Chatzinotas A, Gallien L, Gobet A, Kurm V, Küsel K, Rillig MC, Rivett DW, Salles JF, van der Heijden MGA, Youssef NH, Zhang X, Wei Z, Hol WHG. 2017. Where less may be more: how the rare biosphere pulls ecosystems strings. ISME J 11:853–862. doi: 10.1038/ismej.2016.174. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
