

Maternal and zygotic gene regulatory effects of endogenous RNAi pathways
- PMID: 30759082
- PMCID: PMC6391025
- DOI: 10.1371/journal.pgen.1007784
Maternal and zygotic gene regulatory effects of endogenous RNAi pathways
Abstract
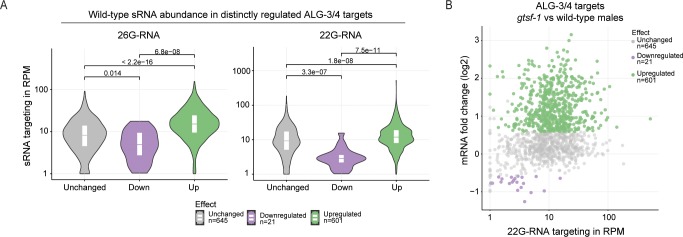
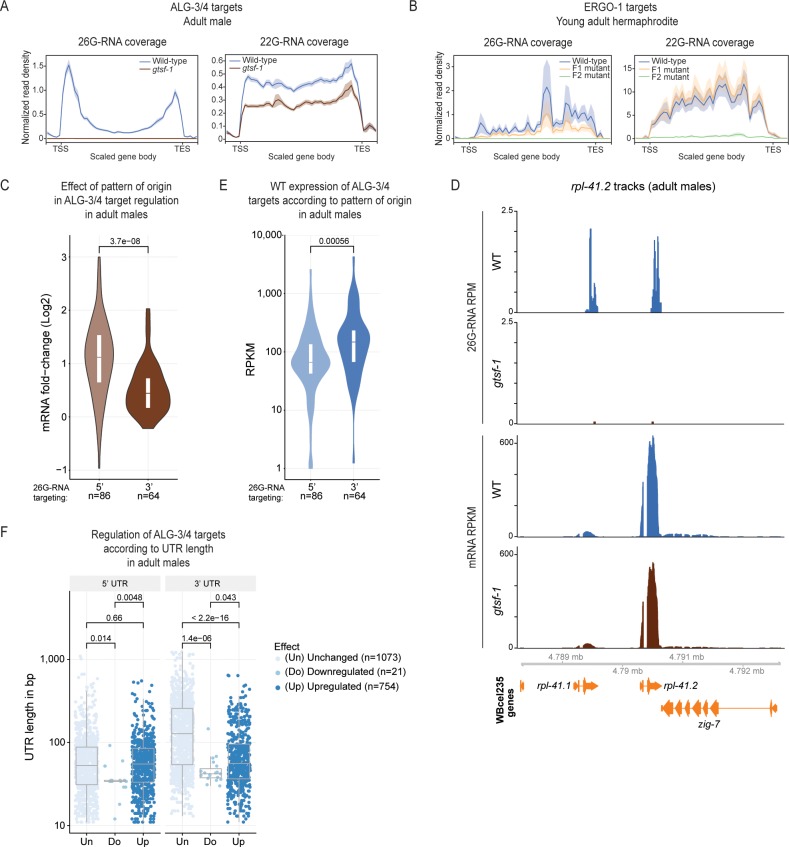
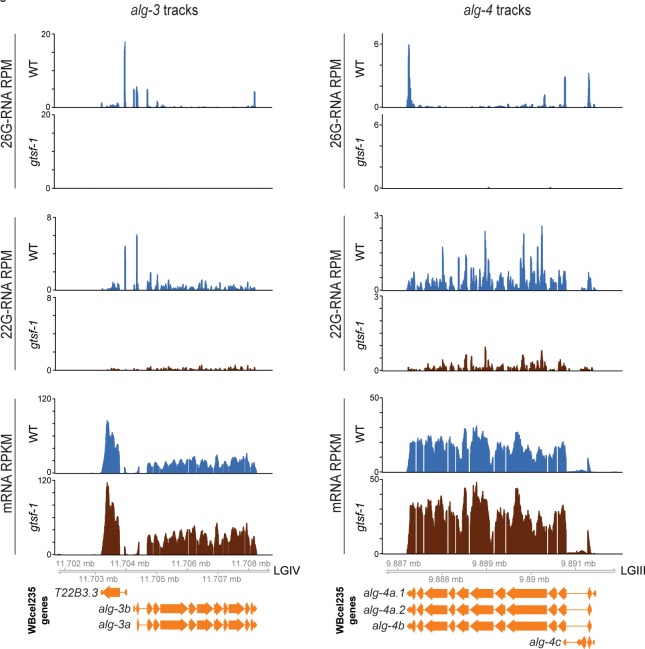
Endogenous small RNAs (sRNAs) and Argonaute proteins are ubiquitous regulators of gene expression in germline and somatic tissues. sRNA-Argonaute complexes are often expressed in gametes and are consequently inherited by the next generation upon fertilization. In Caenorhabditis elegans, 26G-RNAs are primary endogenous sRNAs that trigger the expression of downstream secondary sRNAs. Two subpopulations of 26G-RNAs exist, each of which displaying strongly compartmentalized expression: one is expressed in the spermatogenic gonad and associates with the Argonautes ALG-3/4; plus another expressed in oocytes and in embryos, which associates with the Argonaute ERGO-1. The determinants and dynamics of gene silencing elicited by 26G-RNAs are largely unknown. Here, we provide diverse new insights into these endogenous sRNA pathways of C. elegans. Using genetics and deep sequencing, we dissect a maternal effect of the ERGO-1 branch of the 26G-RNA pathway. We find that maternal primary sRNAs can trigger the production of zygotic secondary sRNAs that are able to silence targets, even in the absence of zygotic primary triggers. Thus, the interaction of maternal and zygotic sRNA populations, assures target gene silencing throughout animal development. Furthermore, we explore other facets of 26G-RNA biology related to the ALG-3/4 branch. We find that sRNA abundance, sRNA pattern of origin and the 3' UTR length of target transcripts are predictors of the regulatory outcome by the Argonautes ALG-3/4. Lastly, we provide evidence suggesting that ALG-3 and ALG-4 regulate their own mRNAs in a negative feedback loop. Altogether, we provide several new regulatory insights on the dynamics, target regulation and self-regulation of the endogenous RNAi pathways of C. elegans.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
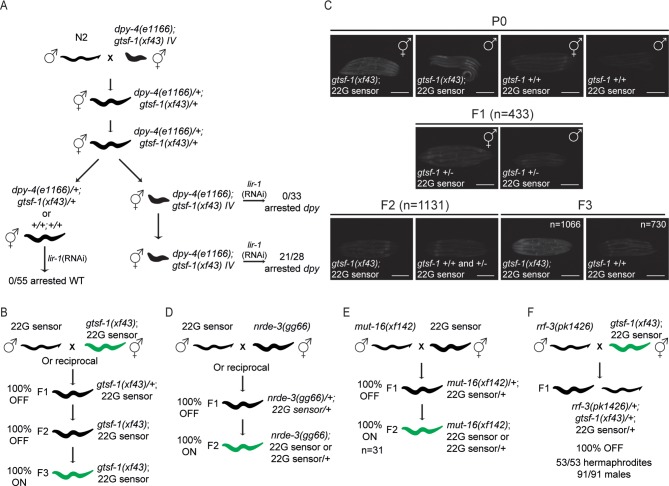
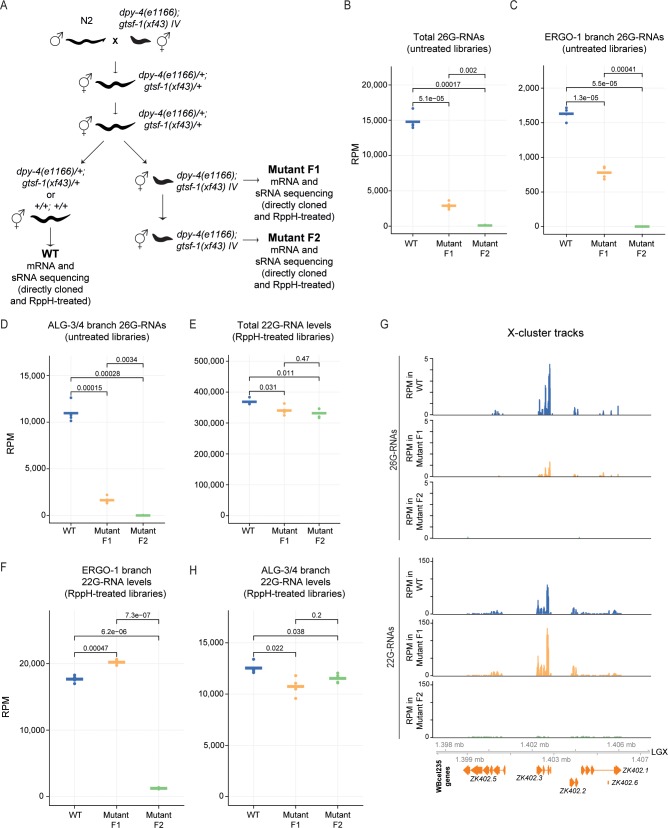
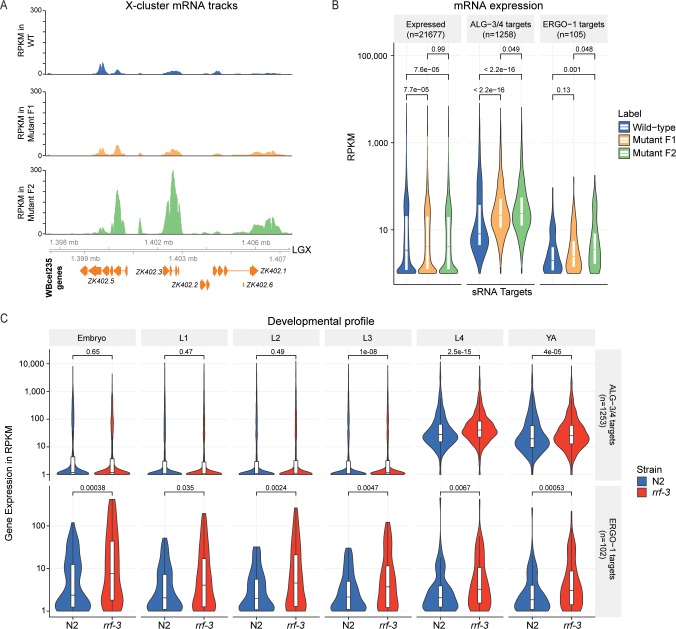
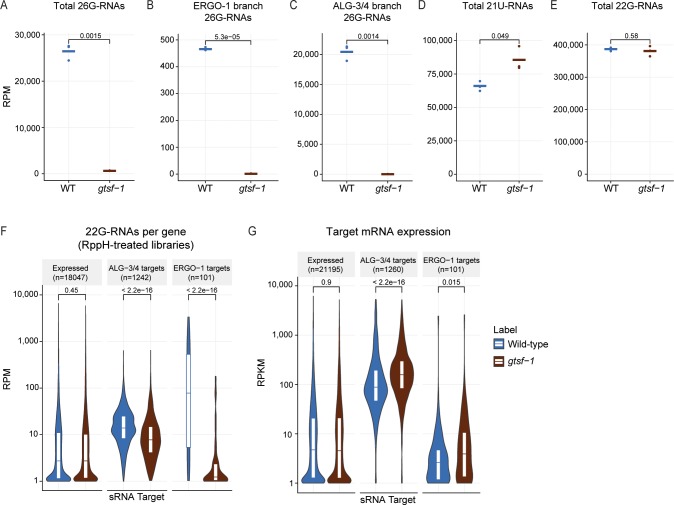
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
