

Dynamic inosinome profiles reveal novel patient stratification and gender-specific differences in glioblastoma
- PMID: 30760294
- PMCID: PMC6373152
- DOI: 10.1186/s13059-019-1647-x
Dynamic inosinome profiles reveal novel patient stratification and gender-specific differences in glioblastoma
Abstract
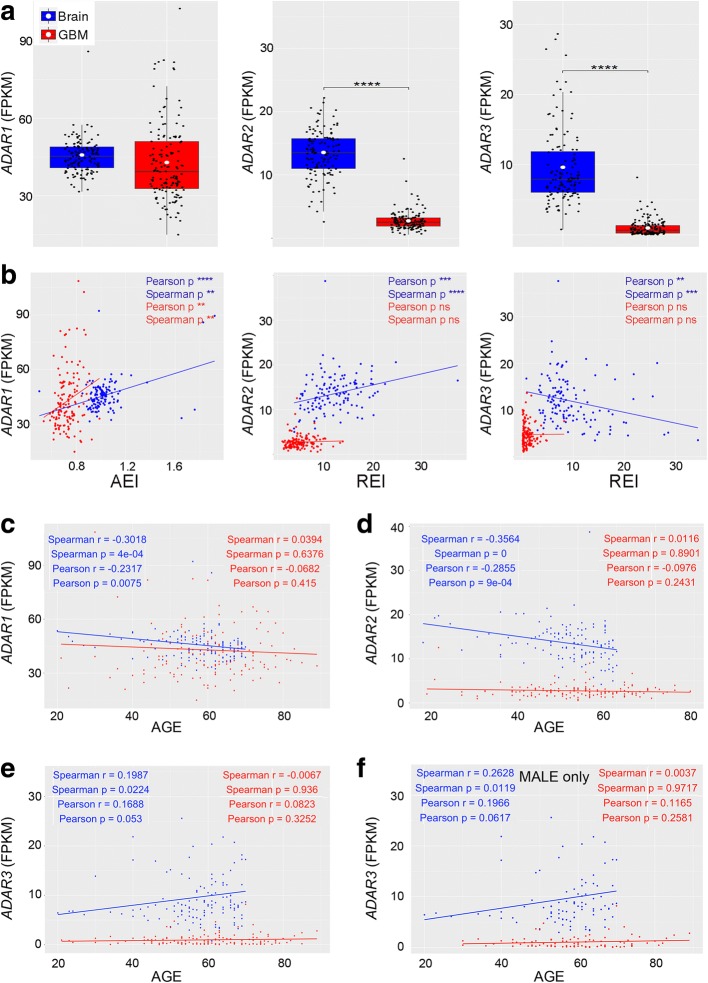
Background: Adenosine-to-inosine (A-to-I) RNA editing is an essential post-transcriptional mechanism mediated by ADAR enzymes that have been recently associated with cancer.
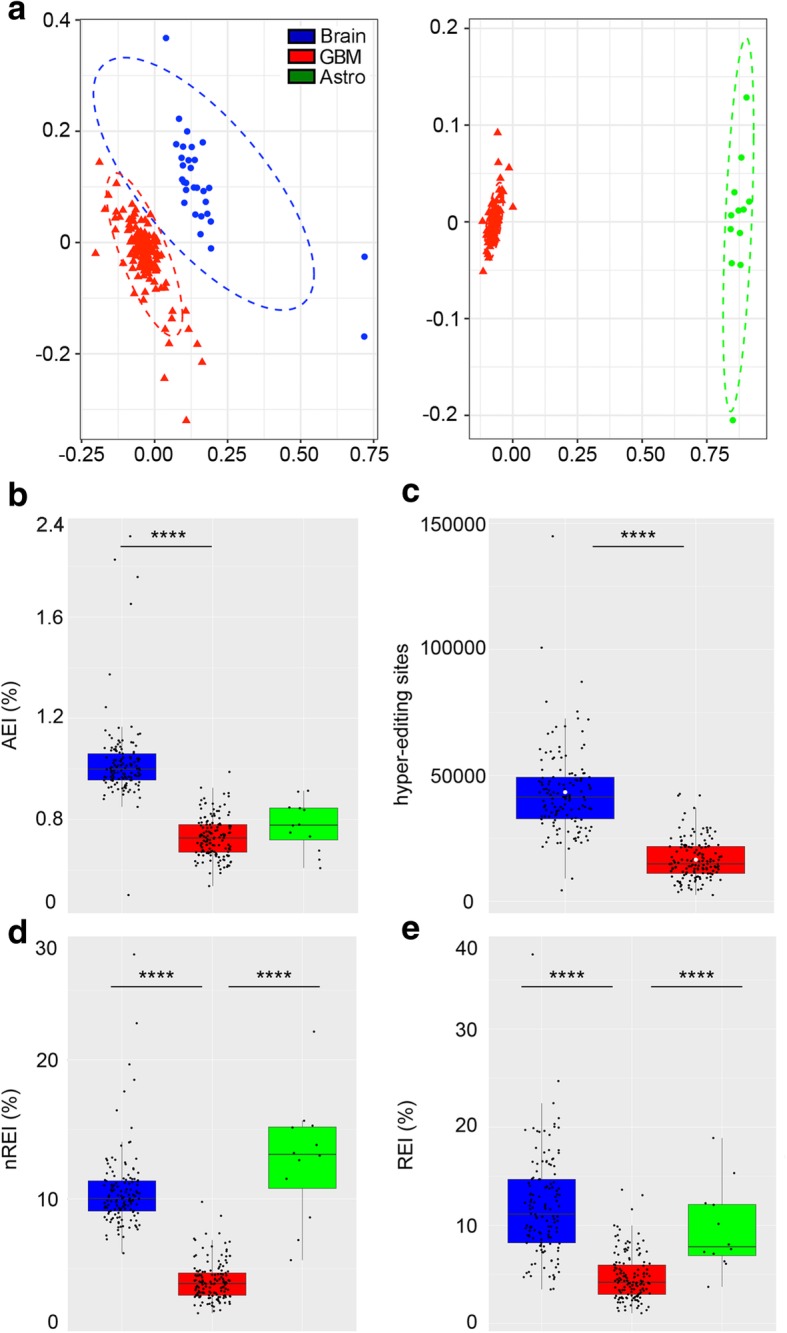
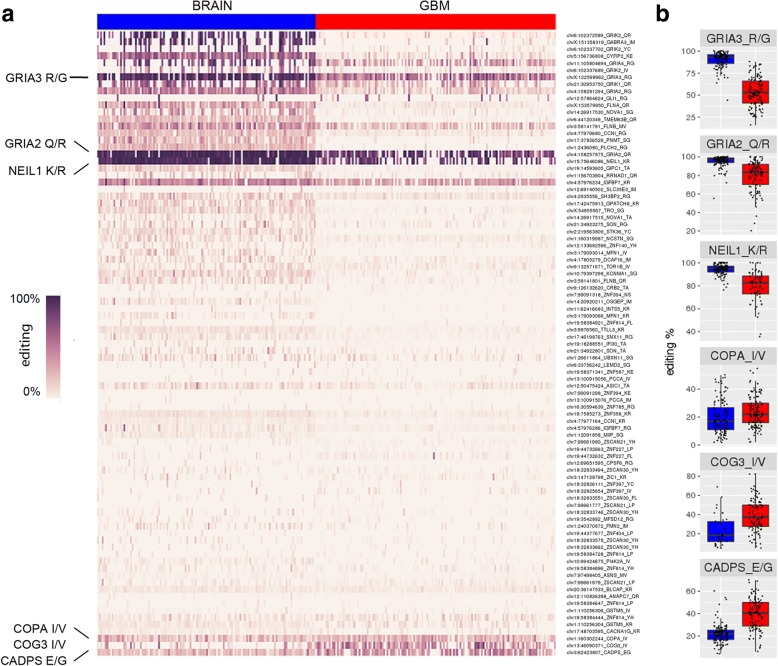
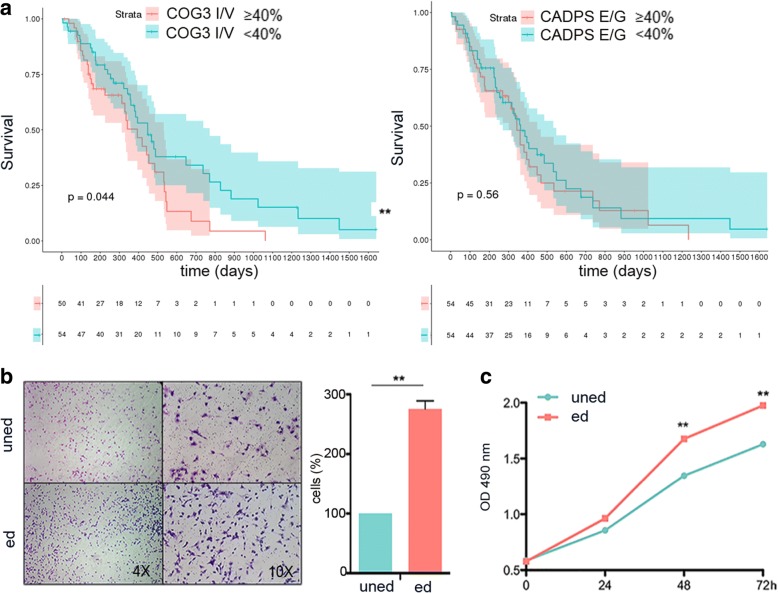
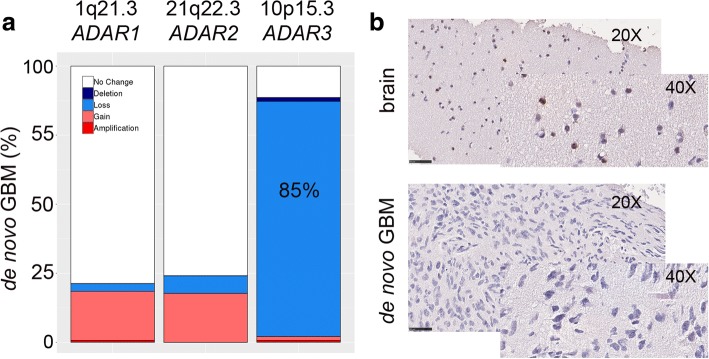
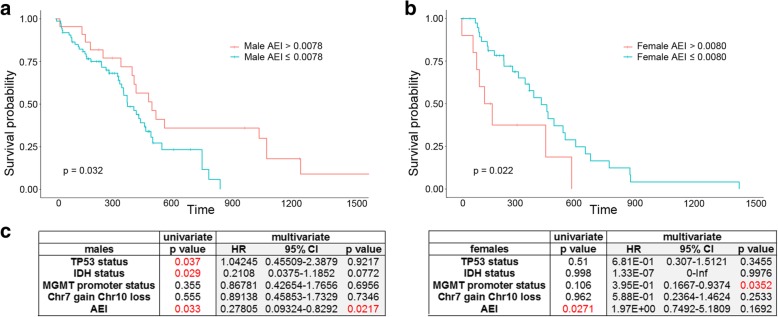
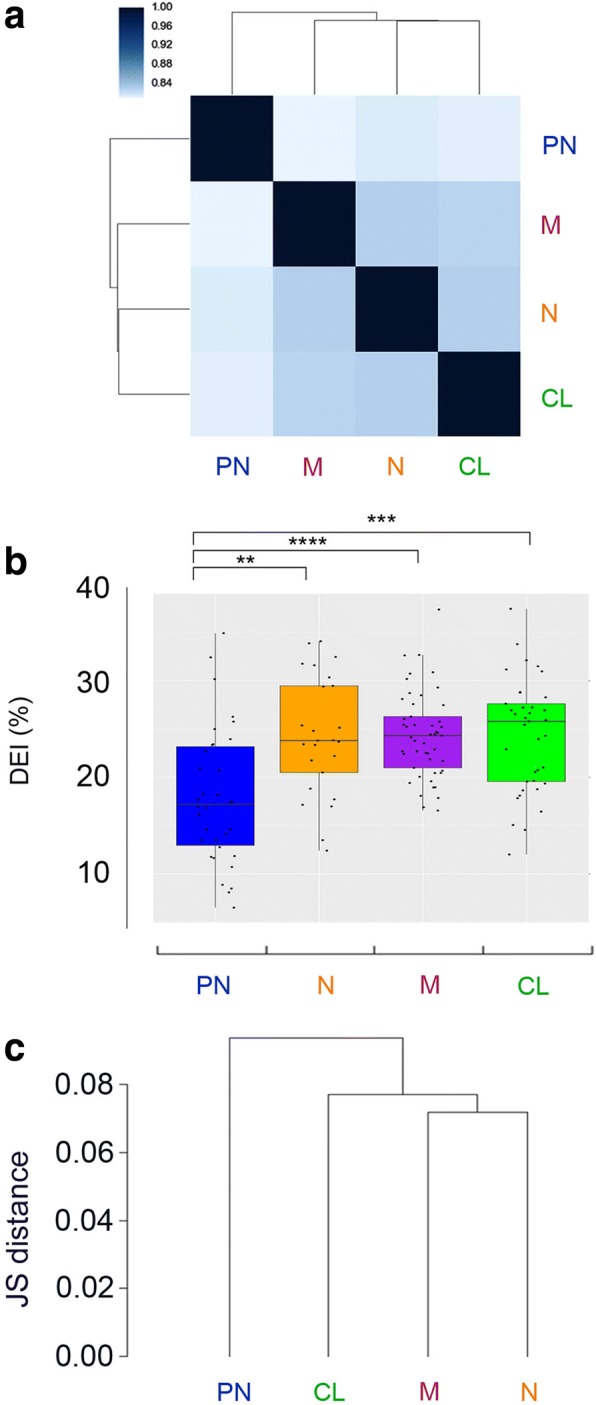
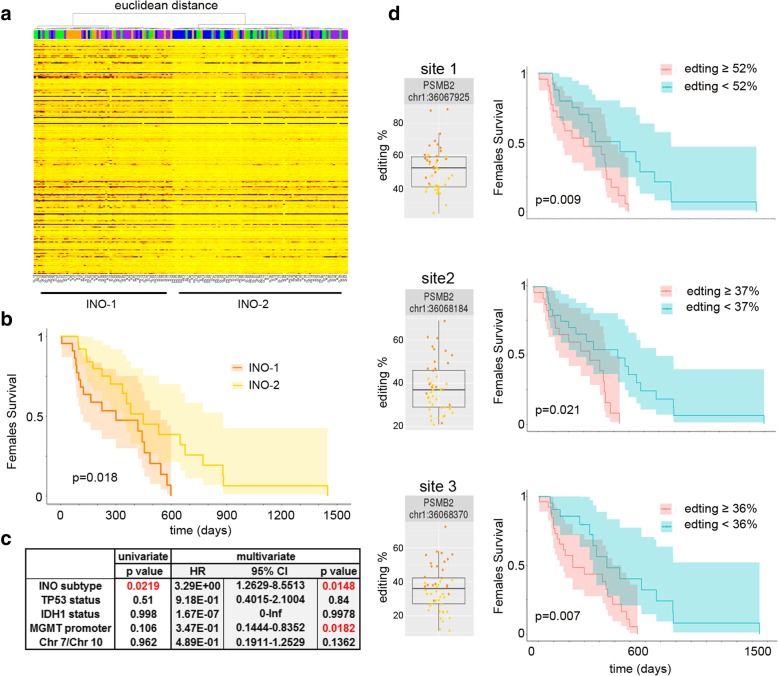
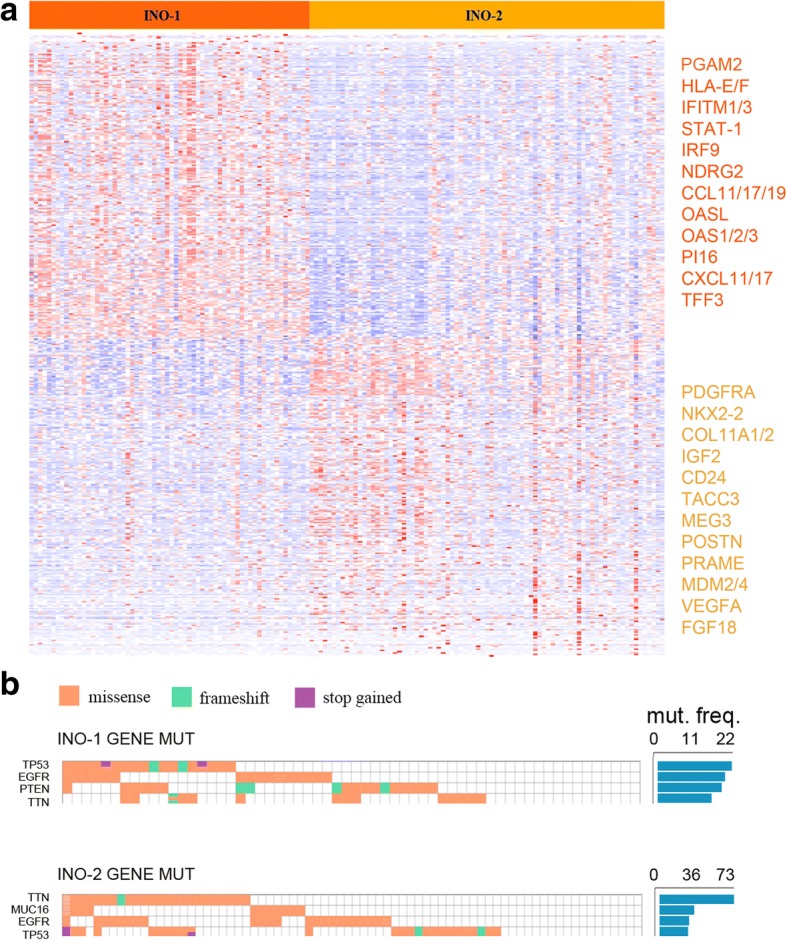
Results: Here, we characterize the inosinome signature in normal brain and de novo glioblastoma (GBM) using new metrics that re-stratify GBM patients according to their editing profiles and indicate this post-transcriptional event as a possible molecular mechanism for sexual dimorphism in GBM. We find that over 85% of de novo GBMs carry a deletion involving the genomic locus of ADAR3, which is specifically expressed in the brain. By analyzing RNA editing and patient outcomes, an intriguing gender-dependent link appears, with high editing of Alus shown to be beneficial only in male patients. We propose an inosinome-based molecular stratification of GBM patients that identifies two different GBM subgroups, INO-1 and INO-2, which can identify novel high-risk gender-specific patient groups for which more aggressive treatments may be necessary.
Conclusions: Our data provide a detailed picture of RNA editing landscape in normal brain and GBM, exploring A-to-I RNA editing regulation, disclosing unexpected editing implications for GBM patient stratification and identification of gender-dependent high-risk patients, and suggesting COG3 I/V as an eligible site for future personalized targeted gene therapy.
Keywords: ADAR; COG3; GBM; RNA editing; RNA-Seq.
Conflict of interest statement
Ethics approval and consent to participate
The study was approved by the local committee on the use of human samples for experimental studies of the Catholic University School of Medicine, Rome, Italy (Prot. 12045/15). Written informed consents were provided by the participants prior to enrollment. All experimental methods abided by the Helsinki Declaration.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
