

Longitudinal HIV sequencing reveals reservoir expression leading to decay which is obscured by clonal expansion
- PMID: 30760706
- PMCID: PMC6374386
- DOI: 10.1038/s41467-019-08431-7
Longitudinal HIV sequencing reveals reservoir expression leading to decay which is obscured by clonal expansion
Abstract
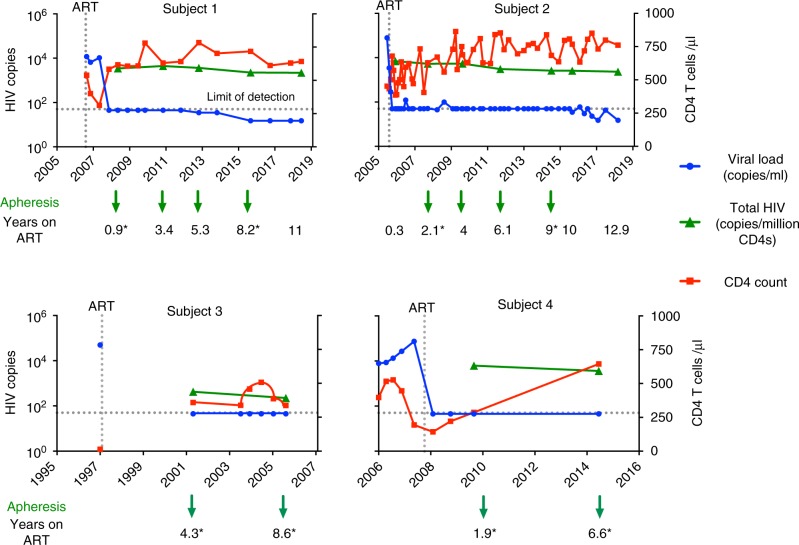
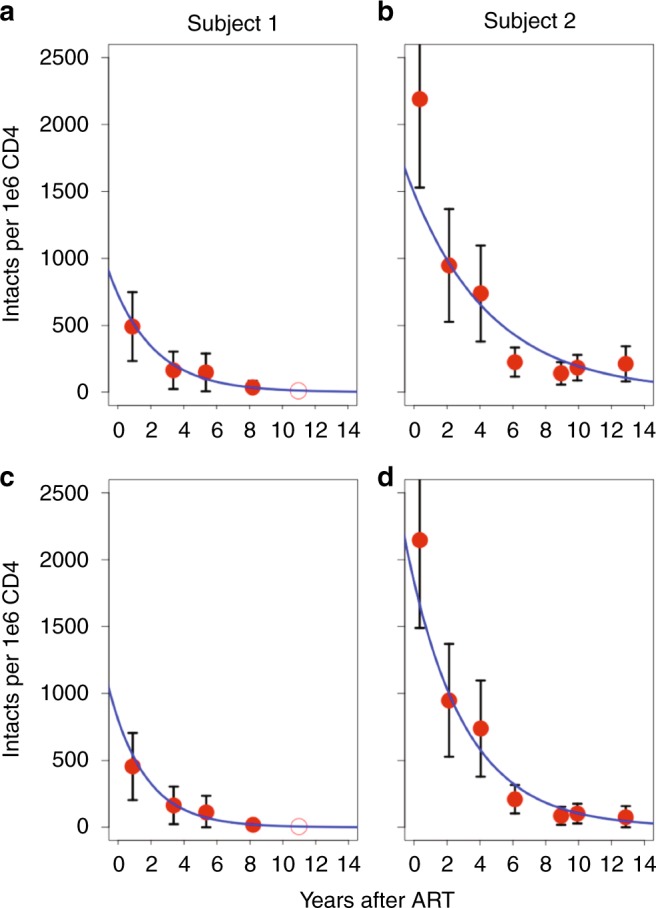
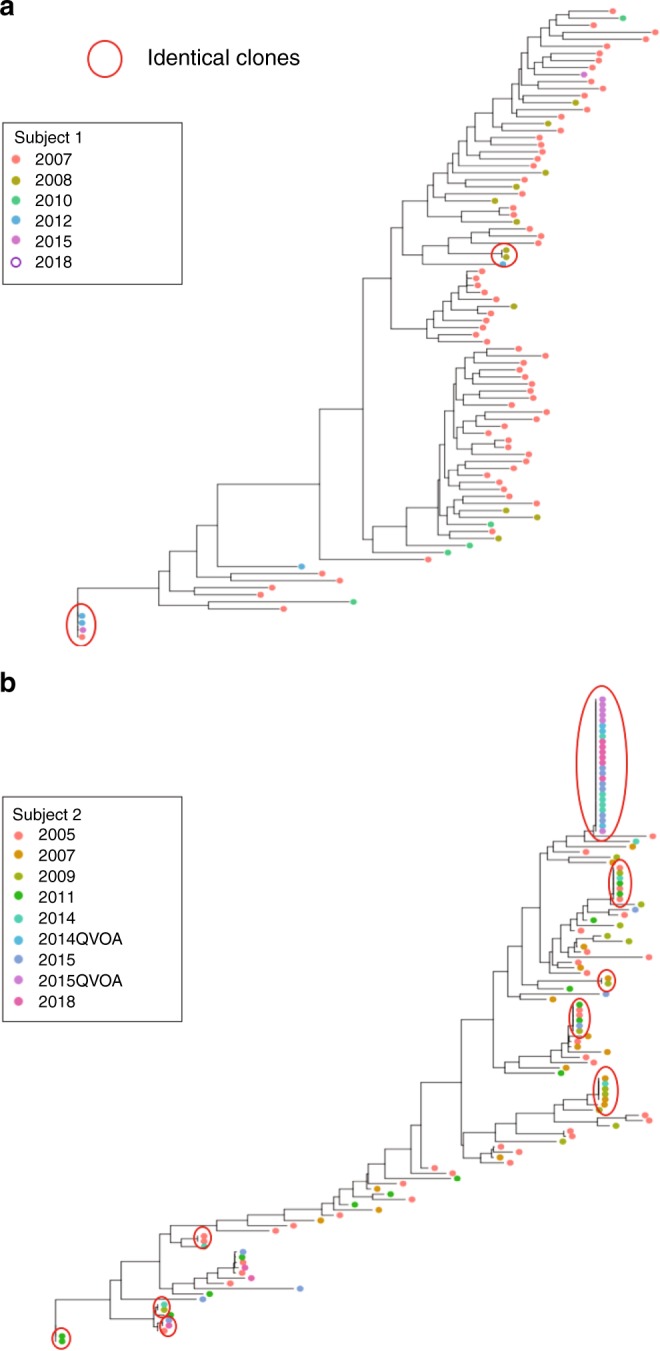
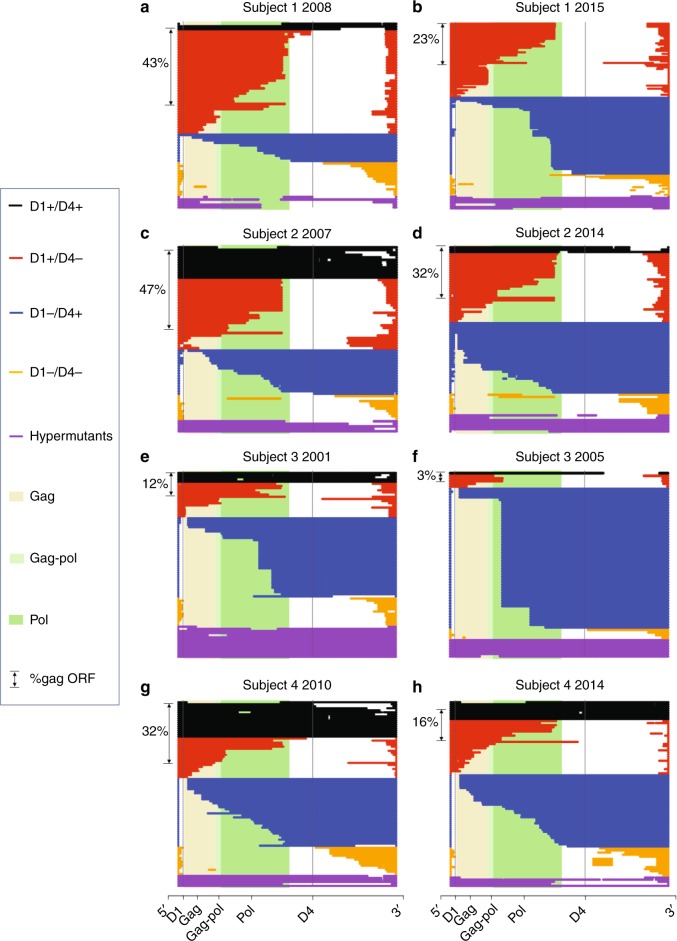
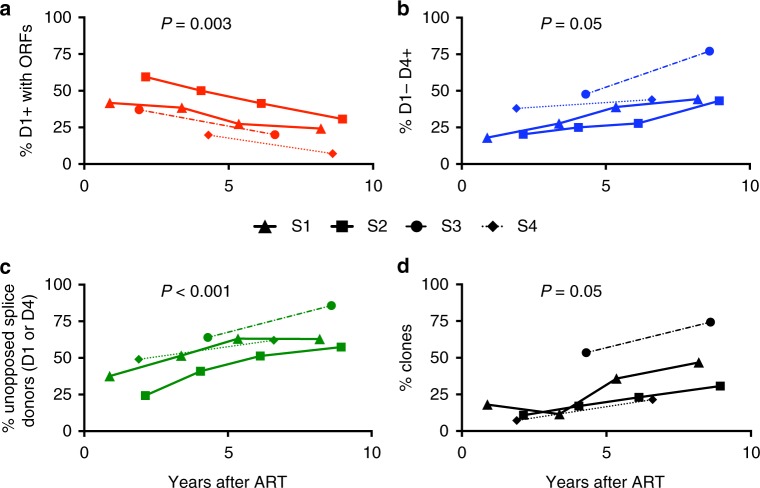
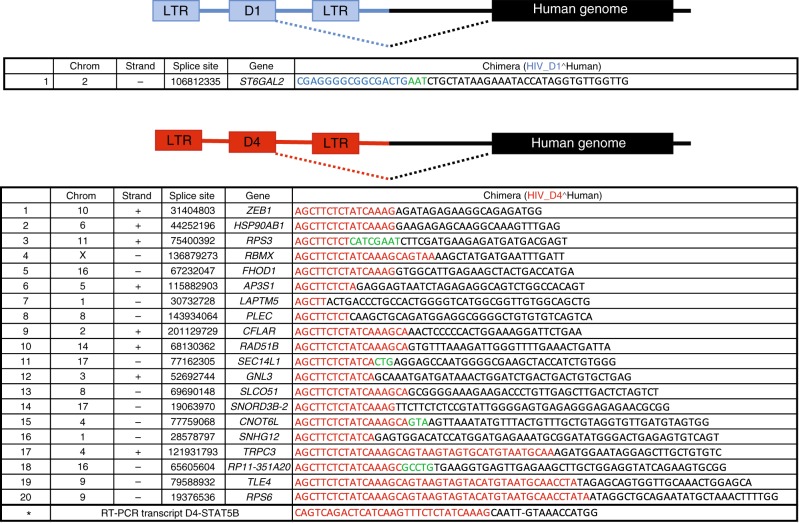
After initiating antiretroviral therapy (ART), a rapid decline in HIV viral load is followed by a long period of undetectable viremia. Viral outgrowth assay suggests the reservoir continues to decline slowly. Here, we use full-length sequencing to longitudinally study the proviral landscape of four subjects on ART to investigate the selective pressures influencing the dynamics of the treatment-resistant HIV reservoir. We find intact and defective proviruses that contain genetic elements favoring efficient protein expression decrease over time. Moreover, proviruses that lack these genetic elements, yet contain strong donor splice sequences, increase relatively to other defective proviruses, especially among clones. Our work suggests that HIV expression occurs to a significant extent during ART and results in HIV clearance, but this is obscured by the expansion of proviral clones. Paradoxically, clonal expansion may also be enhanced by HIV expression that leads to splicing between HIV donor splice sites and downstream human exons.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R21 AI143564/AI/NIAID NIH HHS/United States
- R33 AI104280/AI/NIAID NIH HHS/United States
- R21 AI116216/AI/NIAID NIH HHS/United States
- R21AI116216/NH/NIH HHS/United States
- UM1 AI126617/AI/NIAID NIH HHS/United States
- UM1AI126617/NH/NIH HHS/United States
- UM1 AI126619/AI/NIAID NIH HHS/United States
- R01 AI120011/AI/NIAID NIH HHS/United States
- R01-AI140970 /NH/NIH HHS/United States
- R01AI12001 /NH/NIH HHS/United States
- R01 AI140970/AI/NIAID NIH HHS/United States
- R33AI104280/NH/NIH HHS/United States
- R37 AI150556/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
