

Transferrin receptor 1 is required for enucleation of mouse erythroblasts during terminal differentiation
- PMID: 30761254
- PMCID: PMC6356176
- DOI: 10.1002/2211-5463.12573
Transferrin receptor 1 is required for enucleation of mouse erythroblasts during terminal differentiation
Abstract
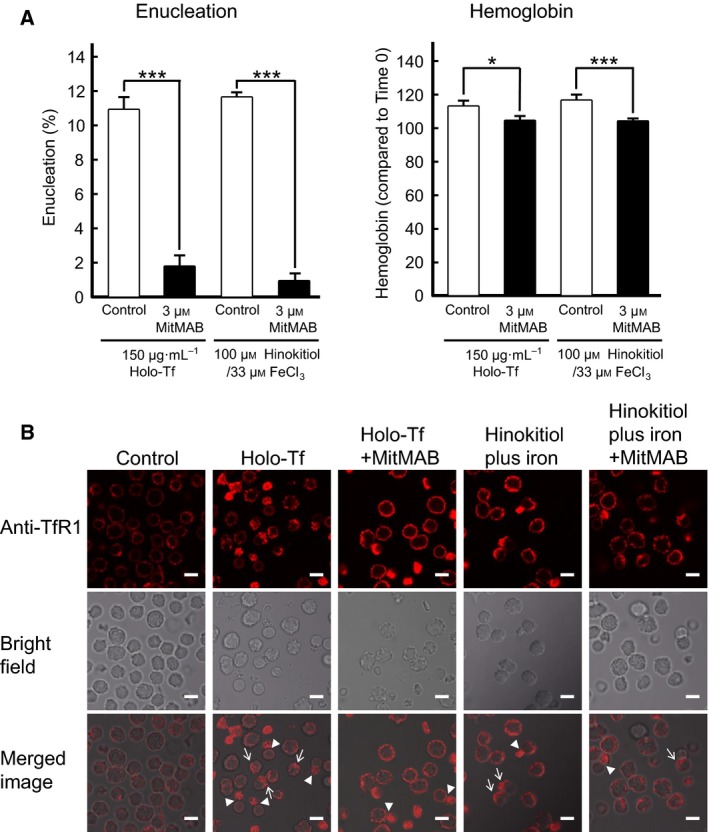
Enucleation is the process whereby the nucleus is extruded from the erythroblast during late stage mammalian erythropoiesis. However, the specific signaling pathways involved in this process remain unclear. To better understand the mechanisms underlying erythroblast enucleation, we investigated erythroblast enucleation using both the spleens of adult mice with phenylhydrazine-induced anemia and mouse fetal livers. Our results indicated that both iron-bound transferrin (holo-Tf) and the small-molecule iron transporter hinokitiol with iron ions (hinokitiol plus iron) promote hemoglobin synthesis and the enucleation of mouse spleen-derived erythroblasts. Although an antitransferrin receptor 1 (TfR1) monoclonal antibody inhibited both enucleation and hemoglobin synthesis promoted by holo-Tf, it inhibited only enucleation, but not hemoglobin synthesis, promoted by hinokitiol plus iron. Furthermore, siRNA against mouse TfR1 were found to suppress the enucleation of mouse fetal liver-derived erythroblasts, and the endocytosis inhibitor MitMAB inhibited enucleation, hemoglobin synthesis, and the internalization of TfR1 promoted by both types of stimuli. Collectively, our results suggest that TfR1, iron ions, and endocytosis play important roles in mouse erythroblast enucleation.
Keywords: enucleation; erythroblast; hinokitiol; iron; transferrin; transferrin receptor.
Figures




Similar articles
-
Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts.Haematologica. 2010 Dec;95(12):2013-21. doi: 10.3324/haematol.2010.029827. Epub 2010 Sep 7. Haematologica. 2010. PMID: 20823130 Free PMC article.
-
Erythroblast differentiation at spleen in Q137E mutant ribosomal protein S19 gene knock-in C57BL/6J mice.Immunobiology. 2018 Jan;223(1):118-124. doi: 10.1016/j.imbio.2017.10.003. Epub 2017 Oct 5. Immunobiology. 2018. PMID: 29017823
-
Iron dose-dependent differentiation and enucleation of human erythroblasts in serum-free medium.J Tissue Eng Regen Med. 2016 Feb;10(2):E84-9. doi: 10.1002/term.1743. Epub 2013 Apr 18. J Tissue Eng Regen Med. 2016. PMID: 23606586 Free PMC article.
-
Transferrin Receptors in Erythropoiesis.Int J Mol Sci. 2020 Dec 19;21(24):9713. doi: 10.3390/ijms21249713. Int J Mol Sci. 2020. PMID: 33352721 Free PMC article. Review.
-
Erythropoiesis and transferrin receptors.Curr Opin Hematol. 2015 May;22(3):193-8. doi: 10.1097/MOH.0000000000000133. Curr Opin Hematol. 2015. PMID: 25767952 Review.
Cited by
-
Multi-pathway oxidative stress amplification via controllably targeted nanomaterials for photoimmunotherapy of tumors.J Nanobiotechnology. 2025 Jan 22;23(1):33. doi: 10.1186/s12951-025-03116-4. J Nanobiotechnology. 2025. PMID: 39844145 Free PMC article.
-
Towards manufactured red blood cells for the treatment of inherited anemia.Haematologica. 2021 Sep 1;106(9):2304-2311. doi: 10.3324/haematol.2020.268847. Haematologica. 2021. PMID: 34042406 Free PMC article. Review.
-
Reticulocyte Maturation.Membranes (Basel). 2022 Mar 10;12(3):311. doi: 10.3390/membranes12030311. Membranes (Basel). 2022. PMID: 35323786 Free PMC article. Review.
-
M-CSF supports medullary erythropoiesis and erythroid iron demand following burn injury through its activity on homeostatic iron recycling.Sci Rep. 2022 Jan 24;12(1):1235. doi: 10.1038/s41598-022-05360-2. Sci Rep. 2022. PMID: 35075211 Free PMC article.
-
The role of erythrocytes and erythroid progenitor cells in tumors.Open Life Sci. 2022 Dec 15;17(1):1641-1656. doi: 10.1515/biol-2022-0102. eCollection 2022. Open Life Sci. 2022. PMID: 36567722 Free PMC article. Review.
References
-
- Wu H, Liu X, Jaenisch R and Lodish HF (1995) Generation of committed erythroid BFU‐E and CFU‐E progenitors does not require erythropoietin or the erythropoietin receptor. Cell 83, 59–67. - PubMed
-
- Koury MJ and Bondurant MC (1988) Maintenance by erythropoietin of viability and maturation of murine erythroid precursor cells. J Cell Physiol 137, 65–74. - PubMed
-
- Ji P, Jayapal SR and Lodish HF (2008) Enucleation of cultured mouse fetal erythroblasts requires Rac GTPases and mDia2. Nat Cell Biol 10, 314–321. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
