

Cyclin-dependent kinase 21 is a novel regulator of proliferation and meiosis in the male germline of zebrafish
- PMID: 30763280
- PMCID: PMC6489137
- DOI: 10.1530/REP-18-0386
Cyclin-dependent kinase 21 is a novel regulator of proliferation and meiosis in the male germline of zebrafish
Abstract
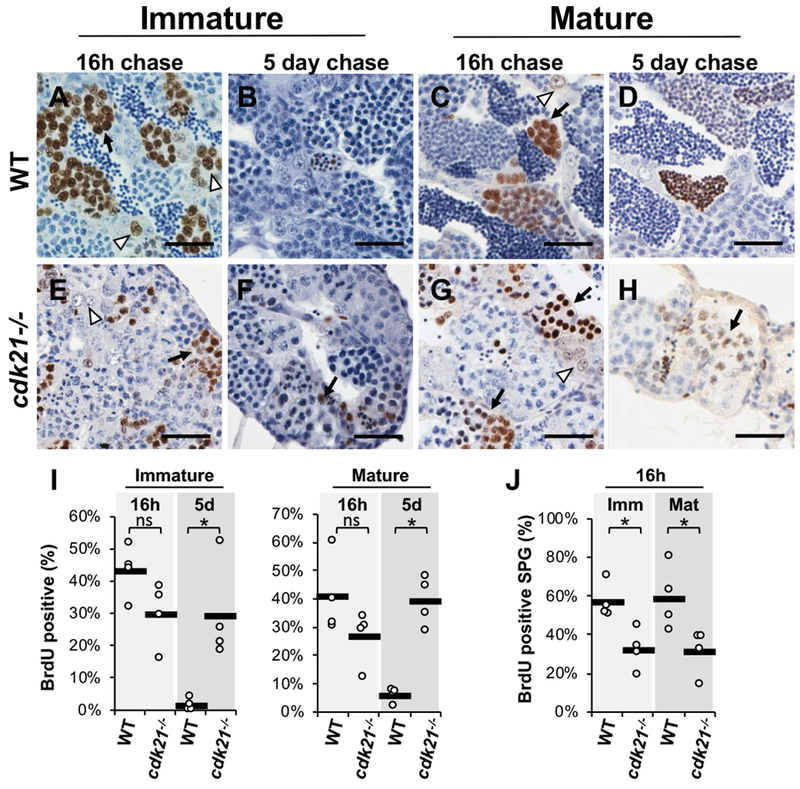
Germ cell differentiation and maintenance relies on complex regulation of mitotic and meiotic progression. Cyclin-dependent kinases (CDKs) and their activating cyclin partners are known to have specialized roles in regulating cell cycle progression across tissues, including germ cells. Very little is known about CDK/cyclin function in zebrafish or the regulation of germ cell maintenance and differentiation. In a forward genetic screen for gonadogenesis defects in zebrafish, a mutation disrupting cdk21 (cyclin-dependent kinase 21) was identified, which caused gonad hypoplasia, reduced fertility and failure of female sex specification. The cdk21 gene is unique to fishes, though the encoded protein is related to the D-cyclin partners Cdk4 and Cdk6, which are known G1 cell cycle regulators. In the testis, cdk21 mutant germ cells exhibited cell cycle defects such as diminished proliferation, prolonged meiosis and delayed sperm differentiation. Furthermore, cdk21 mutants failed to maintain germ cells following breeding. Based on these findings, we propose that cdk21 regulates spermatogonial proliferation, progression through meiosis and germline stem cell activation in the testis. In addition, we investigated cdk4 and cdk6 in zebrafish development and found that each has distinct expression patterns in the gonads. Mutant analysis demonstrated that cdk6 was necessary for viability beyond larval stages. In contrast, cdk4 mutants were viable but were all male with low breeding success and sperm overabundance. Our analysis demonstrated that zebrafish harbor three genes of the cdk4/6 family, cdk4, cdk6 and cdk21, with cdk21 having an essential role in germ cell development in the testis.
Conflict of interest statement
Declaration of Interest
The authors have no conflict of interests to declare.
Figures





Similar articles
-
Diverse roles for CDK-associated activity during spermatogenesis.FEBS Lett. 2019 Oct;593(20):2925-2949. doi: 10.1002/1873-3468.13627. Epub 2019 Oct 20. FEBS Lett. 2019. PMID: 31566717 Free PMC article. Review.
-
Functions of cyclins and CDKs in mammalian gametogenesis†.Biol Reprod. 2019 Sep 1;101(3):591-601. doi: 10.1093/biolre/ioz070. Biol Reprod. 2019. PMID: 31078132 Free PMC article. Review.
-
The RNA-binding protein Adad1 is necessary for germ cell maintenance and meiosis in zebrafish.PLoS Genet. 2023 Aug 8;19(8):e1010589. doi: 10.1371/journal.pgen.1010589. eCollection 2023 Aug. PLoS Genet. 2023. PMID: 37552671 Free PMC article.
-
B cell antigen receptor-mediated activation of cyclin-dependent retinoblastoma protein kinases and inhibition by co-cross-linking with Fc gamma receptors.J Immunol. 1999 Sep 15;163(6):3160-8. J Immunol. 1999. PMID: 10477583
-
Differential phosphorylation of T-47D human breast cancer cell substrates by D1-, D3-, E-, and A-type cyclin-CDK complexes.J Biol Chem. 1997 Dec 26;272(52):33327-37. doi: 10.1074/jbc.272.52.33327. J Biol Chem. 1997. PMID: 9407125
Cited by
-
The Zebrafish Meiotic Cohesin Complex Protein Smc1b Is Required for Key Events in Meiotic Prophase I.Front Cell Dev Biol. 2021 Aug 9;9:714245. doi: 10.3389/fcell.2021.714245. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34434933 Free PMC article.
-
Sexual determination in zebrafish.Cell Mol Life Sci. 2021 Dec 22;79(1):8. doi: 10.1007/s00018-021-04066-4. Cell Mol Life Sci. 2021. PMID: 34936027 Free PMC article. Review.
-
Evolution of the Cdk4/6-Cdkn2 system in invertebrates.Genes Cells. 2024 Nov;29(11):1037-1051. doi: 10.1111/gtc.13165. Epub 2024 Oct 8. Genes Cells. 2024. PMID: 39380239 Free PMC article.
-
Targeting CDK9 for Anti-Cancer Therapeutics.Cancers (Basel). 2021 May 1;13(9):2181. doi: 10.3390/cancers13092181. Cancers (Basel). 2021. PMID: 34062779 Free PMC article. Review.
-
Live and Time-Lapse Imaging of Early Oogenesis and Meiotic Chromosomal Dynamics in Cultured Juvenile Zebrafish Ovaries.Methods Mol Biol. 2021;2218:137-155. doi: 10.1007/978-1-0716-0970-5_12. Methods Mol Biol. 2021. PMID: 33606229
References
-
- Ashley T, Walpita D and de Rooij DG (2001) Localization of two mammalian cyclin dependent kinases during mammalian meiosis. Journal of Cell Science 114 685–693. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
