

Rapid turnover of life-cycle-related genes in the brown algae
- PMID: 30764885
- PMCID: PMC6374913
- DOI: 10.1186/s13059-019-1630-6
Rapid turnover of life-cycle-related genes in the brown algae
Erratum in
-
Correction to: Rapid turnover of life-cycle-related genes in the brown algae.Genome Biol. 2019 Feb 22;20(1):44. doi: 10.1186/s13059-019-1657-8. Genome Biol. 2019. PMID: 30795789 Free PMC article.
Abstract
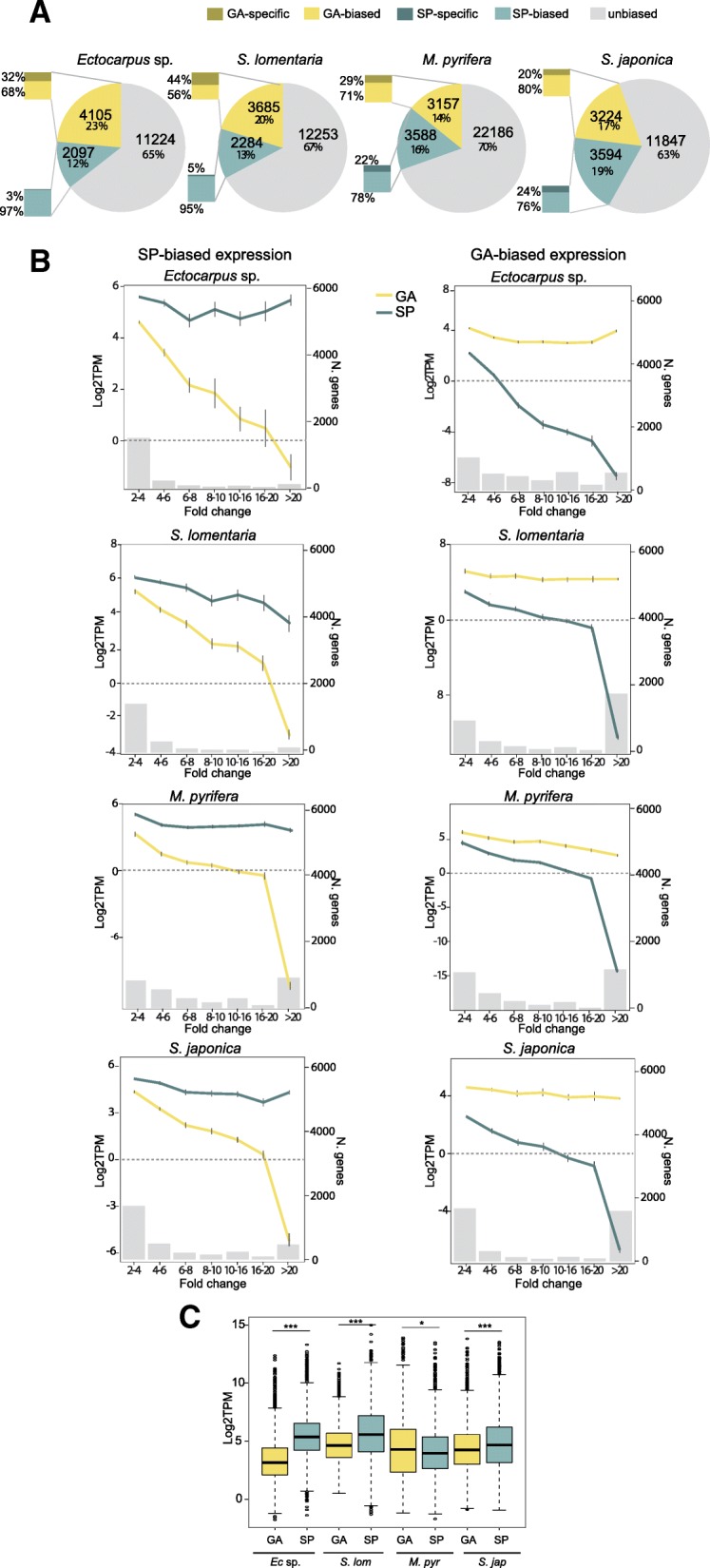
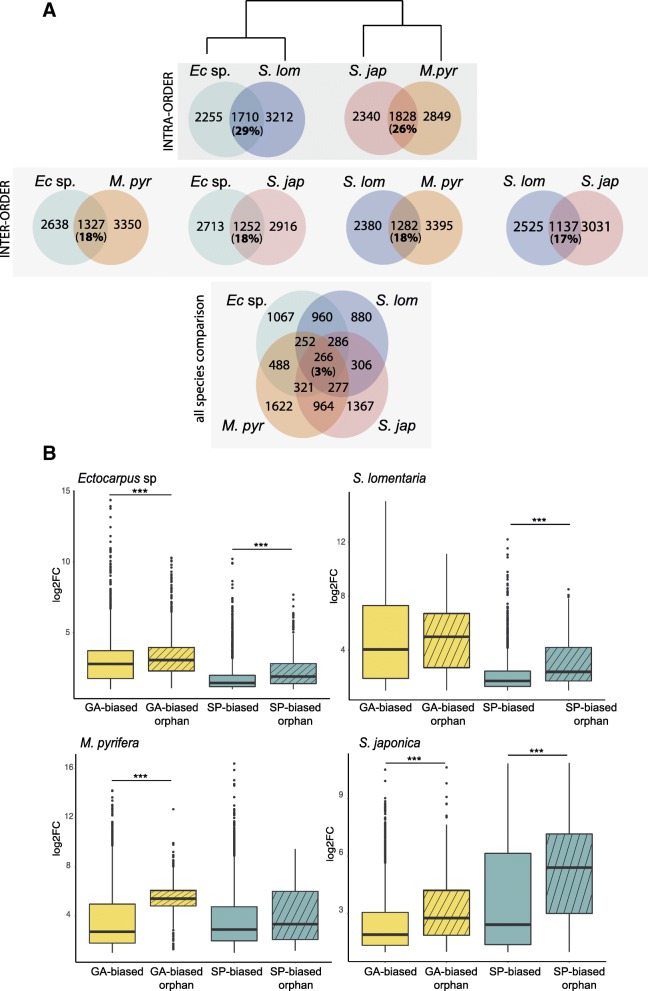
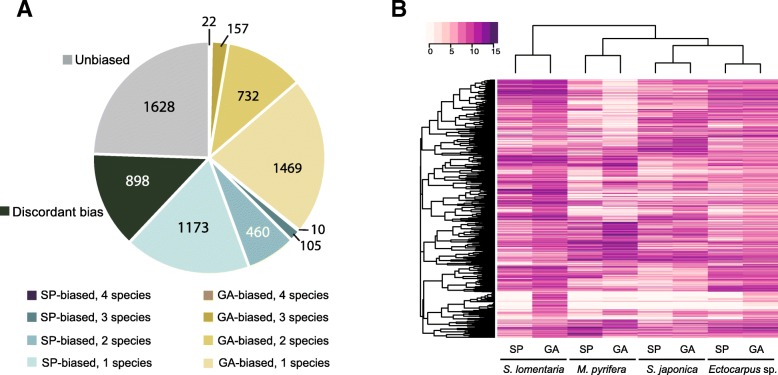
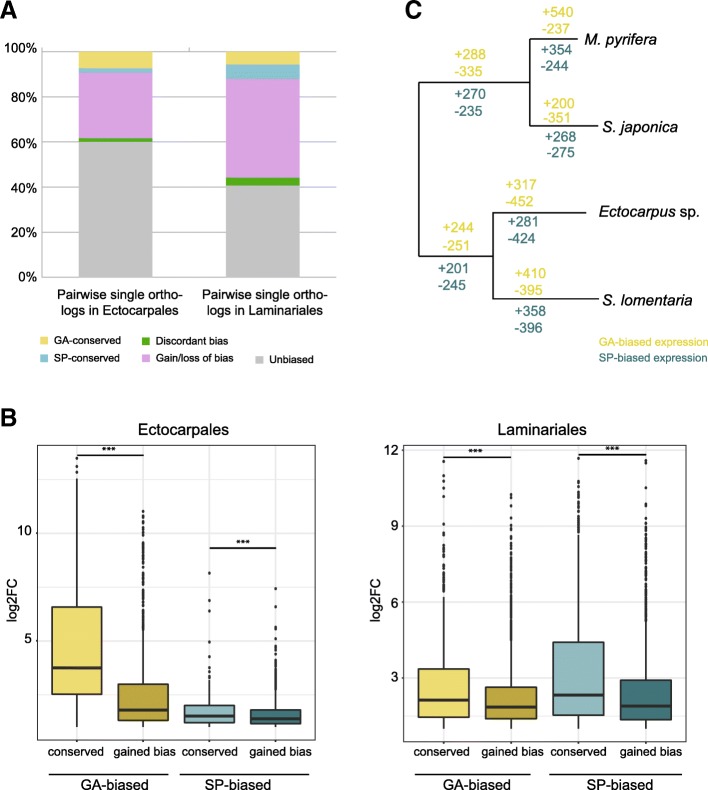
Background: Sexual life cycles in eukaryotes involve a cyclic alternation between haploid and diploid phases. While most animals possess a diploid life cycle, many plants and algae alternate between multicellular haploid (gametophyte) and diploid (sporophyte) generations. In many algae, gametophytes and sporophytes are independent and free-living and may present dramatic phenotypic differences. The same shared genome can therefore be subject to different, even conflicting, selection pressures during each of the life cycle generations. Here, we analyze the nature and extent of genome-wide, generation-biased gene expression in four species of brown algae with contrasting levels of dimorphism between life cycle generations.
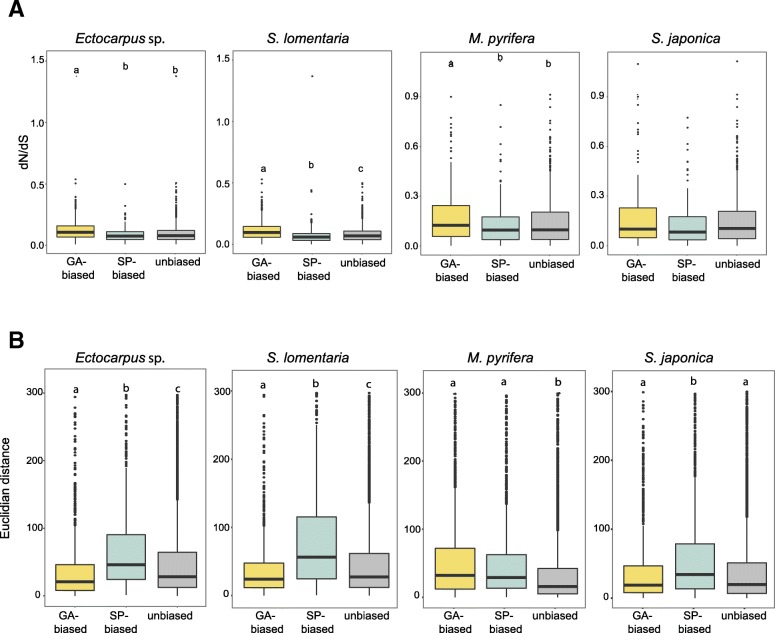
Results: We show that the proportion of the transcriptome that is generation-specific is broadly associated with the level of phenotypic dimorphism between the life cycle stages. Importantly, our data reveals a remarkably high turnover rate for life-cycle-related gene sets across the brown algae and highlights the importance not only of co-option of regulatory programs from one generation to the other but also of a role for newly emerged, lineage-specific gene expression patterns in the evolution of the gametophyte and sporophyte developmental programs in this major eukaryotic group. Moreover, we show that generation-biased genes display distinct evolutionary modes, with gametophyte-biased genes evolving rapidly at the coding sequence level whereas sporophyte-biased genes tend to exhibit changes in their patterns of expression.
Conclusion: Our analysis uncovers the characteristics, expression patterns, and evolution of generation-biased genes and underlines the selective forces that shape this previously underappreciated source of phenotypic diversity.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
