

Emergence of cognitive priming and structure building from the hierarchical interaction of canonical microcircuit models
- PMID: 30767085
- PMCID: PMC6510829
- DOI: 10.1007/s00422-019-00792-y
Emergence of cognitive priming and structure building from the hierarchical interaction of canonical microcircuit models
Abstract
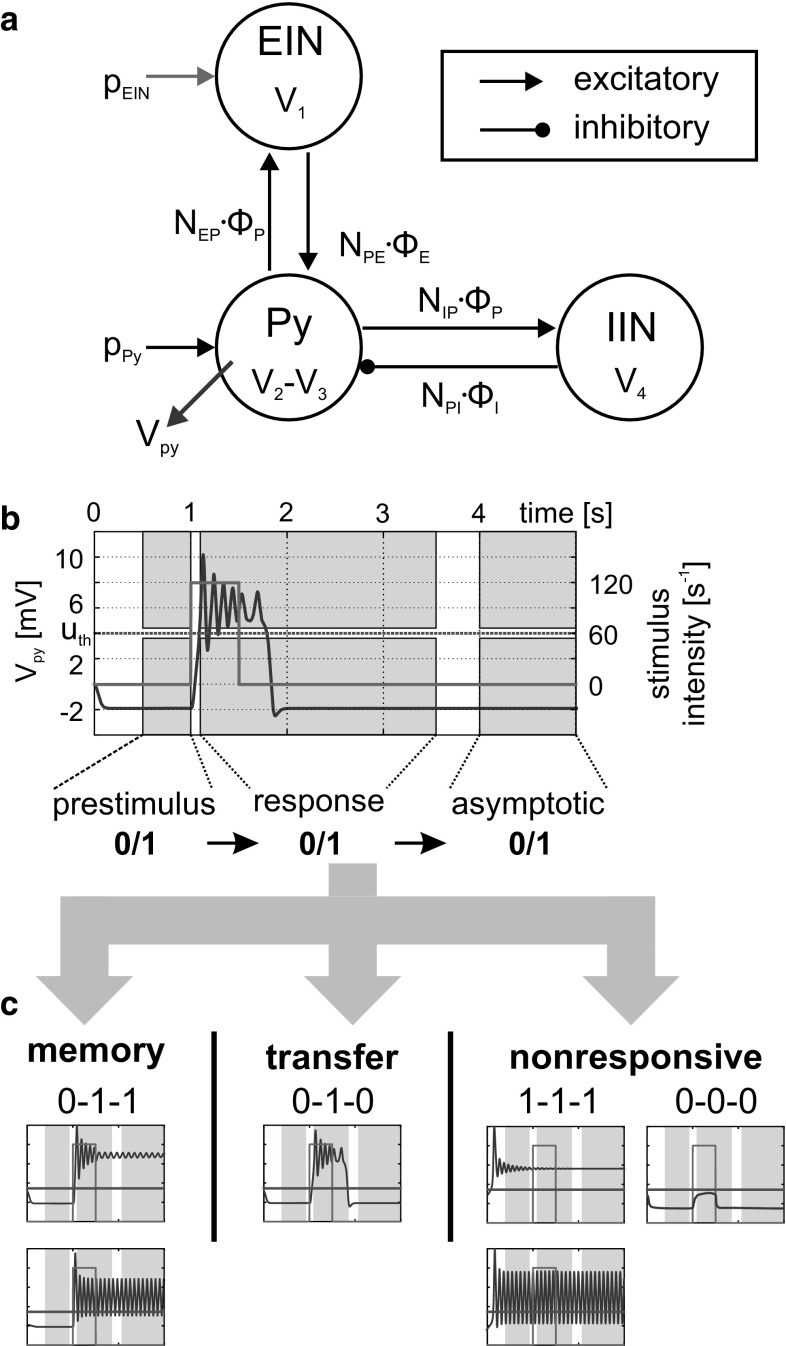
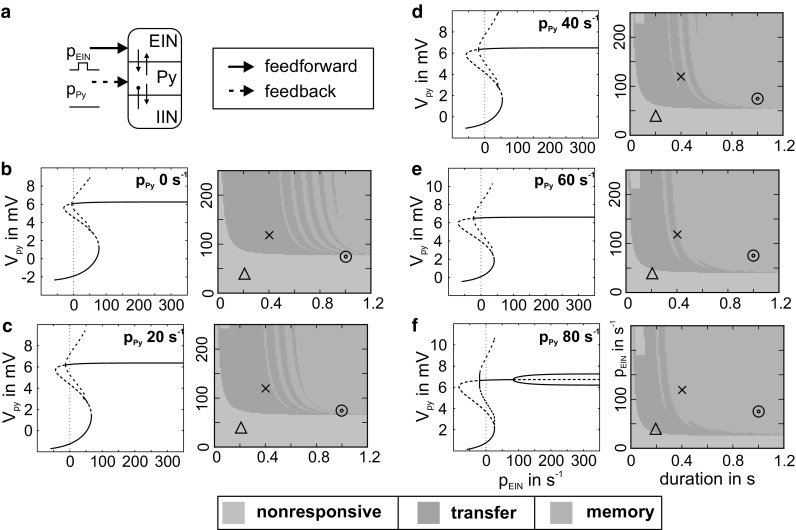
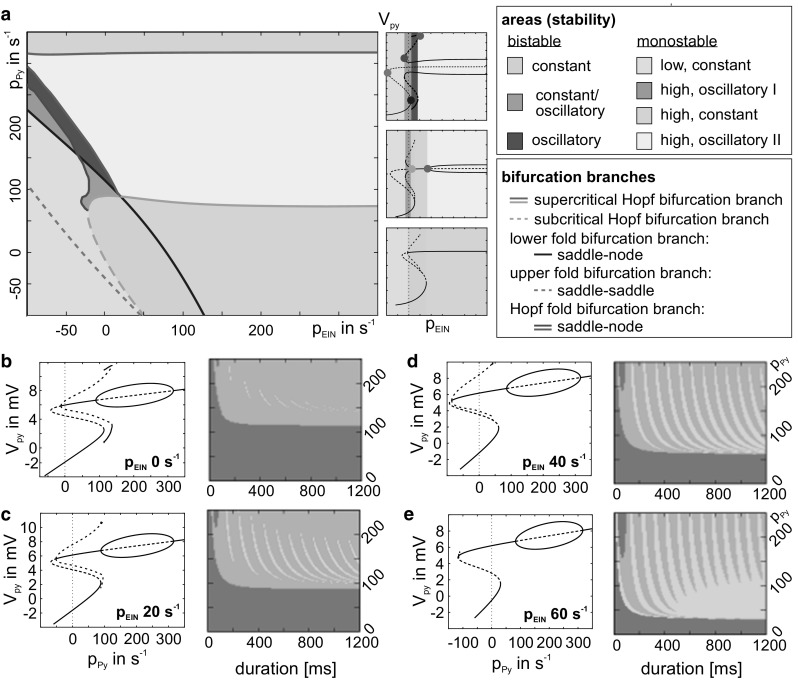
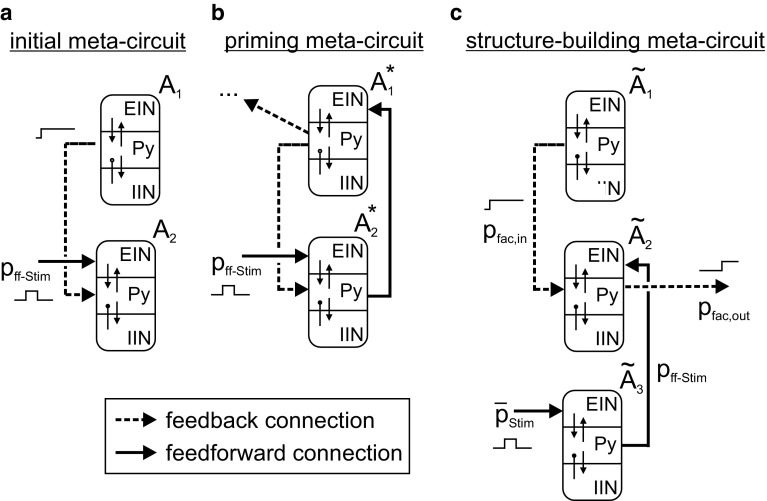
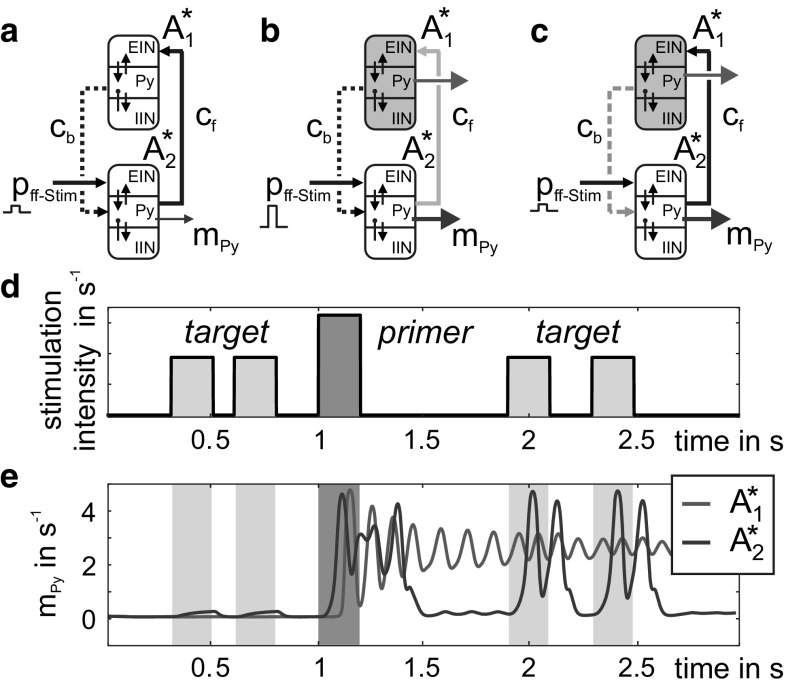
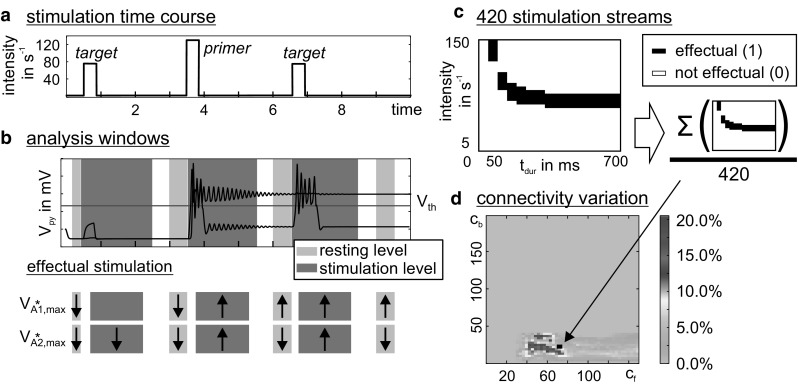
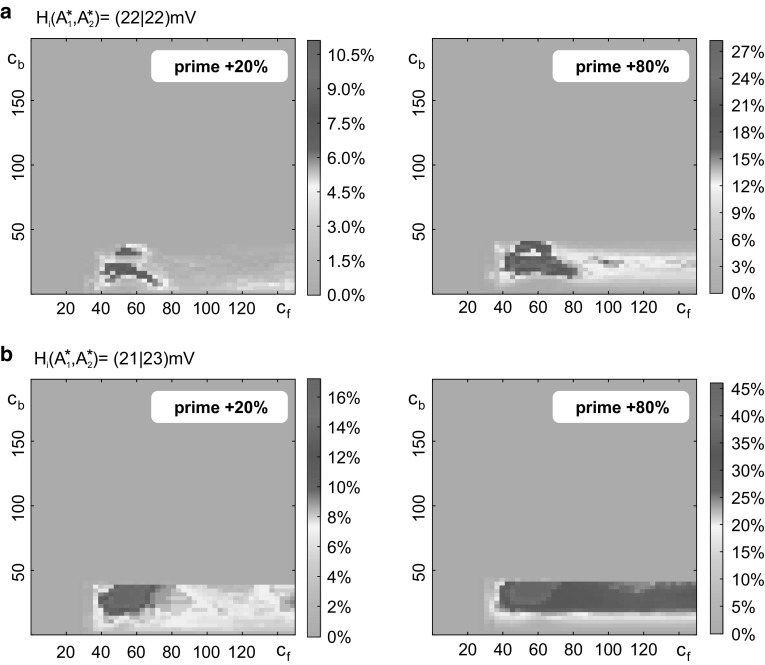
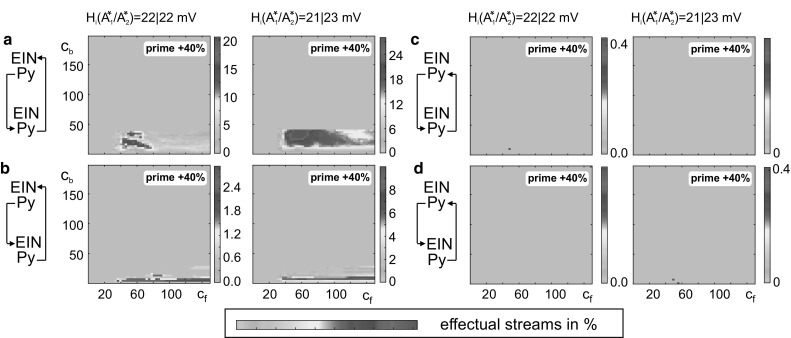
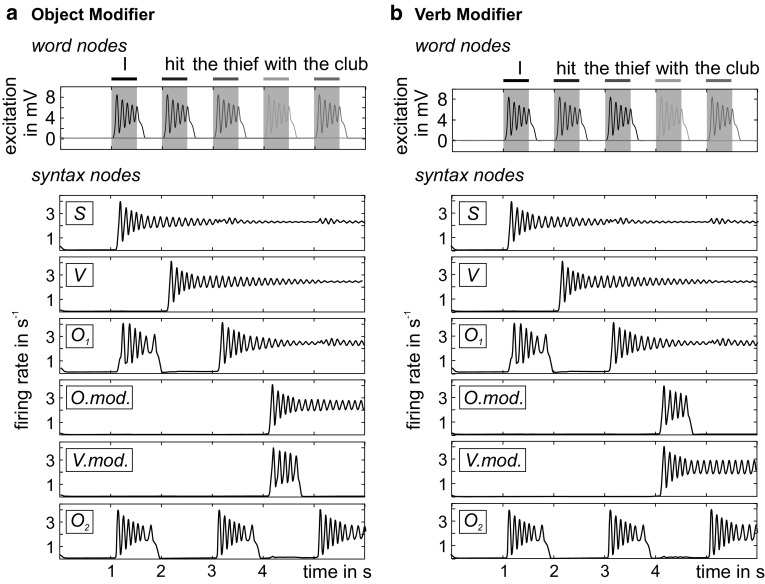
The concept of connectionism states that higher cognitive functions emerge from the interaction of many simple elements. Accordingly, research on canonical microcircuits conceptualizes findings on fundamental neuroanatomical circuits as well as recurrent organizational principles of the cerebral cortex and examines the link between architectures and their associated functionality. In this study, we establish minimal canonical microcircuit models as elements of hierarchical processing networks. Based on a combination of descriptive time simulations and explanatory state-space mappings, we show that minimal canonical microcircuits effectively segregate feedforward and feedback information flows and that feedback information conditions basic processing operations in minimal canonical microcircuits. Further, we derive and examine two prototypical meta-circuits of cooperating minimal canonical microcircuits for the neurocognitive problems of priming and structure building. Through the application of these findings to a language network of syntax parsing, this study embodies neurocognitive research on hierarchical communication in light of canonical microcircuits, cell assembly theory, and predictive coding.
Keywords: Adaptive mechanisms; Canonical microcircuit; Hierarchical model; Neural computations; State-dependent operation; Syntax parsing.
Figures



References
-
- Abeles M. Corticonics, neural circuits of the cerebral cortex. Cambridge: Cambridge University Press; 1991.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
