

Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures
- PMID: 30769894
- PMCID: PMC6406257
- DOI: 10.3390/cells8020160
Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures
Abstract
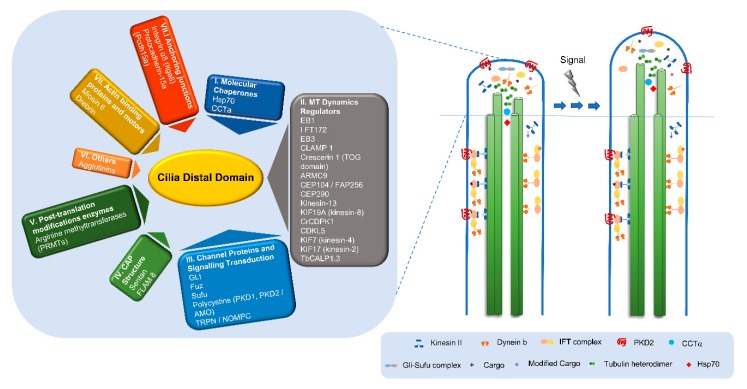
Eukaryotic cilia are microtubule-based organelles that protrude from the cell surface to fulfill sensory and motility functions. Their basic structure consists of an axoneme templated by a centriole/basal body. Striking differences in ciliary ultra-structures can be found at the ciliary base, the axoneme and the tip, not only throughout the eukaryotic tree of life, but within a single organism. Defects in cilia biogenesis and function are at the origin of human ciliopathies. This structural/functional diversity and its relationship with the etiology of these diseases is poorly understood. Some of the important events in cilia function occur at their distal domain, including cilia assembly/disassembly, IFT (intraflagellar transport) complexes' remodeling, and signal detection/transduction. How axonemal microtubules end at this domain varies with distinct cilia types, originating different tip architectures. Additionally, they show a high degree of dynamic behavior and are able to respond to different stimuli. The existence of microtubule-capping structures (caps) in certain types of cilia contributes to this diversity. It has been proposed that caps play a role in axoneme length control and stabilization, but their roles are still poorly understood. Here, we review the current knowledge on cilia structure diversity with a focus on the cilia distal domain and caps and discuss how they affect cilia structure and function.
Keywords: cilia; cilia distal domain; cilia structural diversity; microtubule-capping structures.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
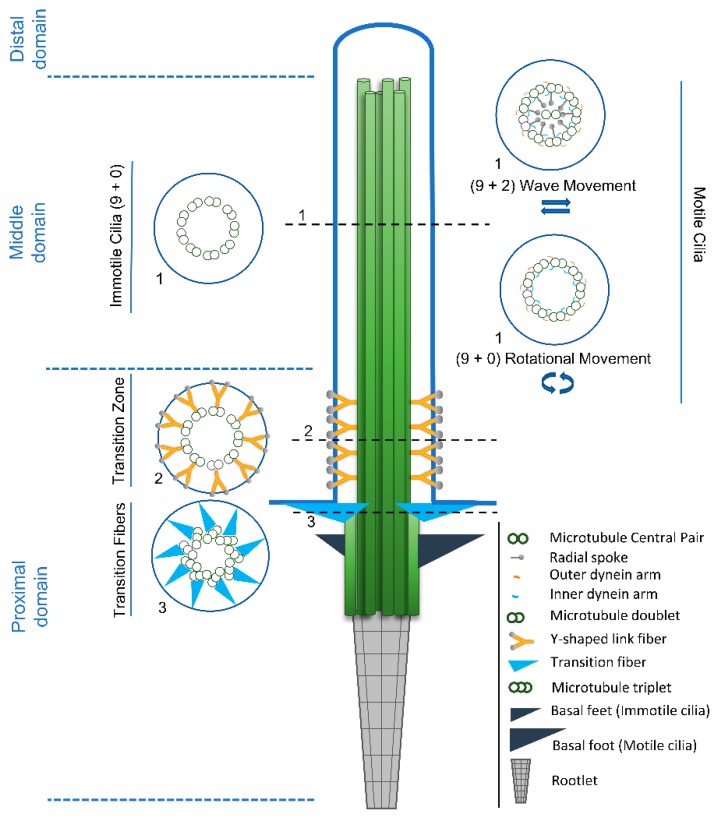
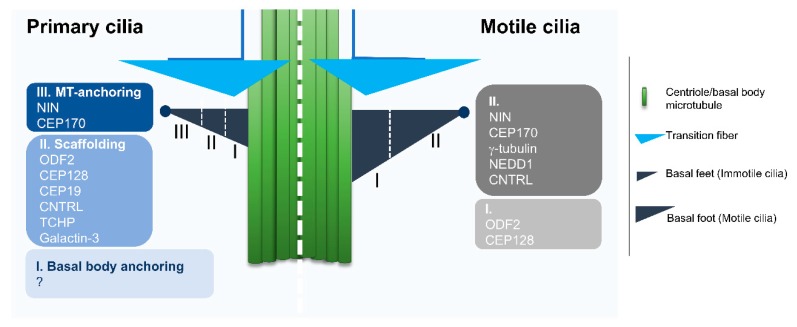
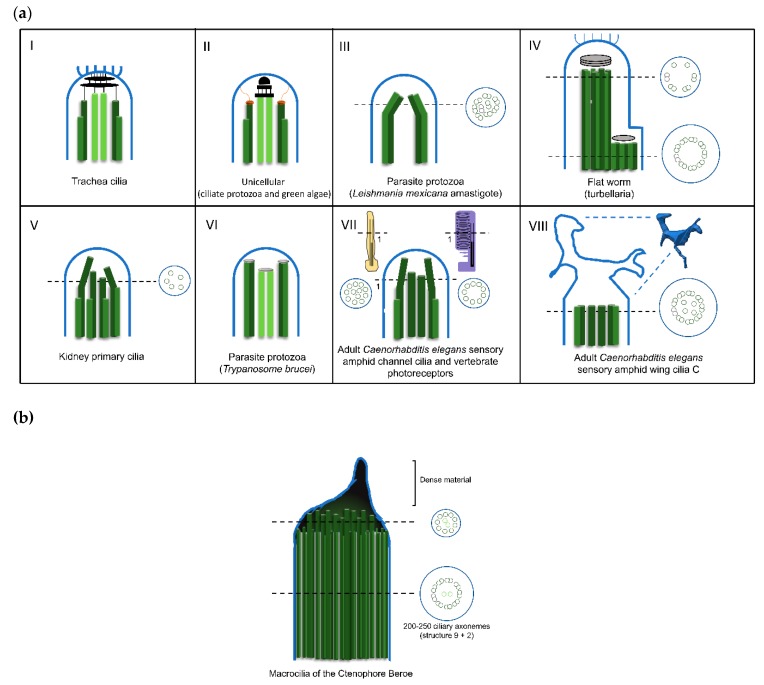
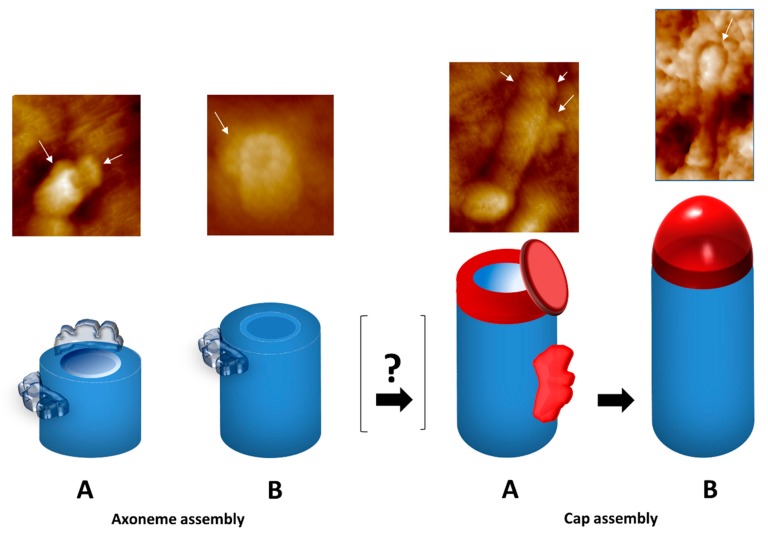
References
-
- Dellinger O.P. The cilium as a key to the structure of contractile protoplasm. J. Morphol. 1909;20:171–210. doi: 10.1002/jmor.1050200202. - DOI
-
- Schmitt F.O., Hall C.E., Jakus M.A. The Ultrastructure of Protoplasmic Fibrils. Biol. Symp. 1943;10:261–276.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
