

A Macrophage-Pericyte Axis Directs Tissue Restoration via Amphiregulin-Induced Transforming Growth Factor Beta Activation
- PMID: 30770250
- PMCID: PMC6436929
- DOI: 10.1016/j.immuni.2019.01.008
A Macrophage-Pericyte Axis Directs Tissue Restoration via Amphiregulin-Induced Transforming Growth Factor Beta Activation
Abstract
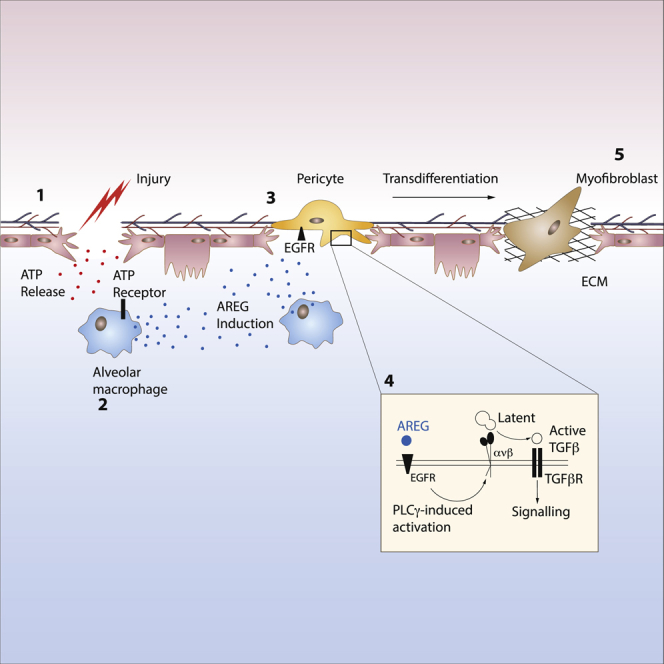
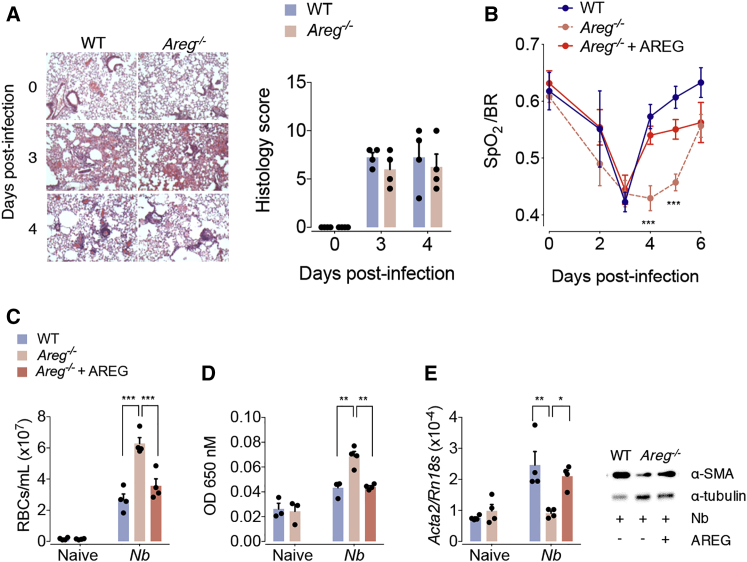
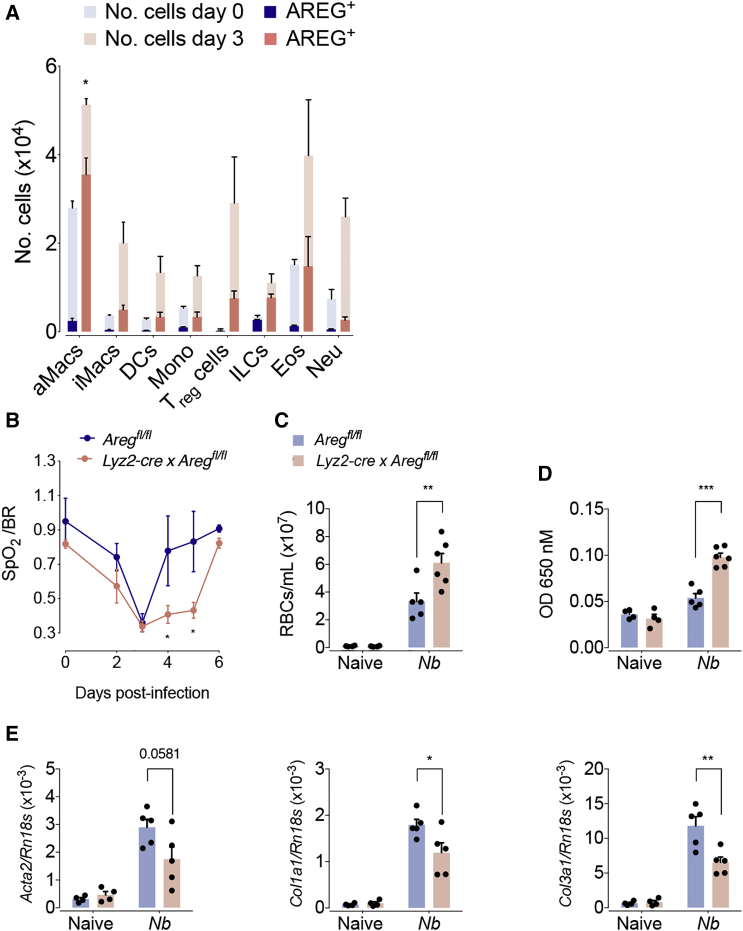
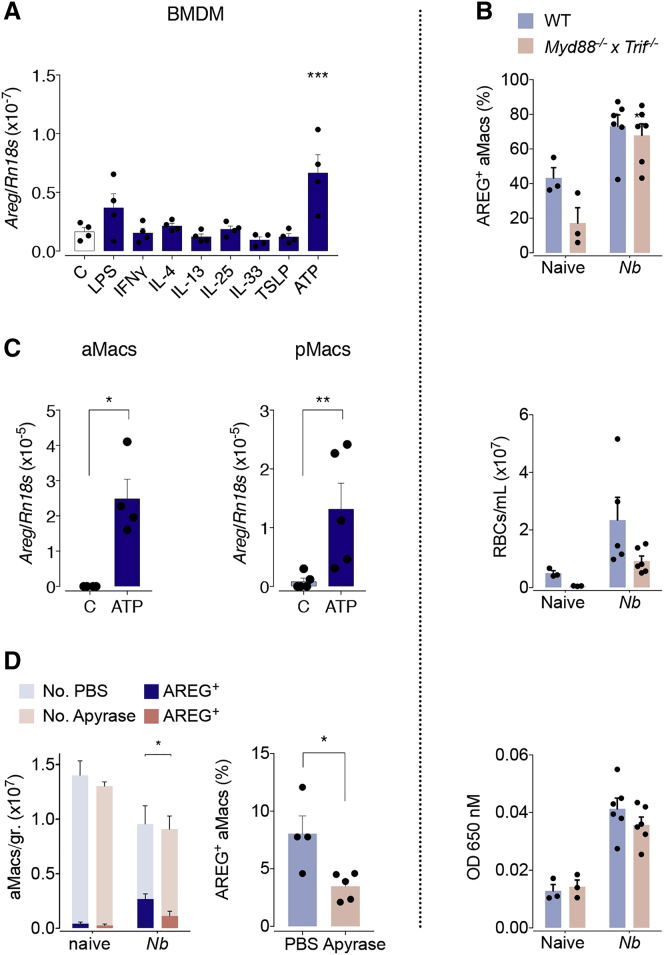
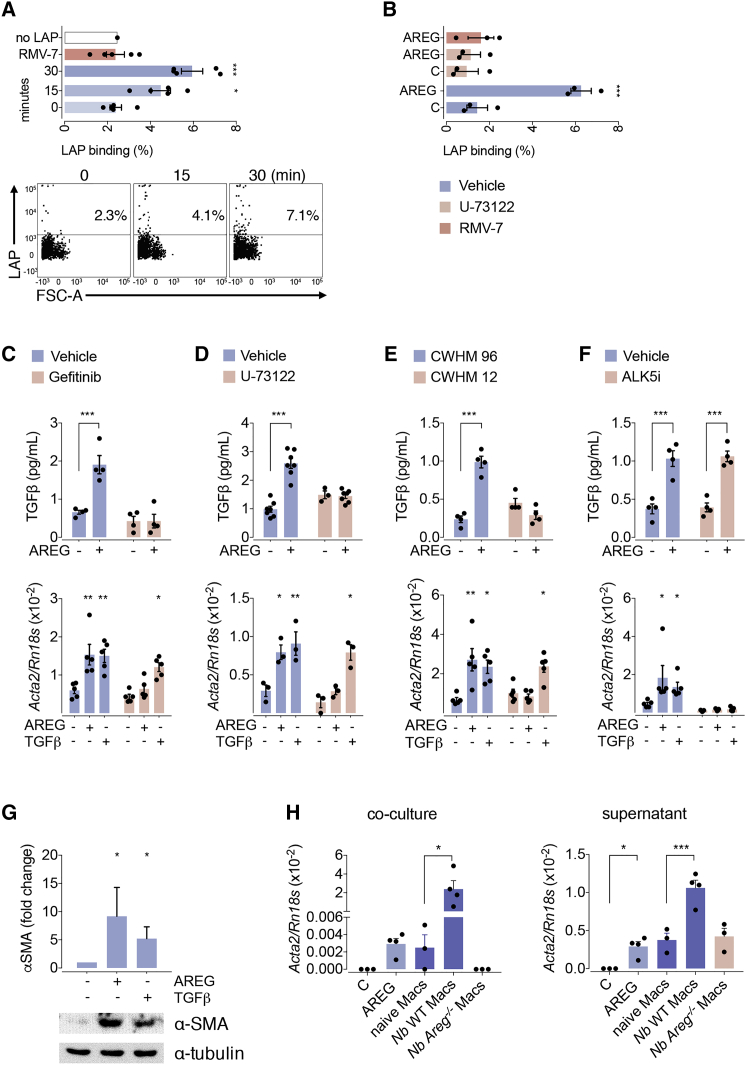
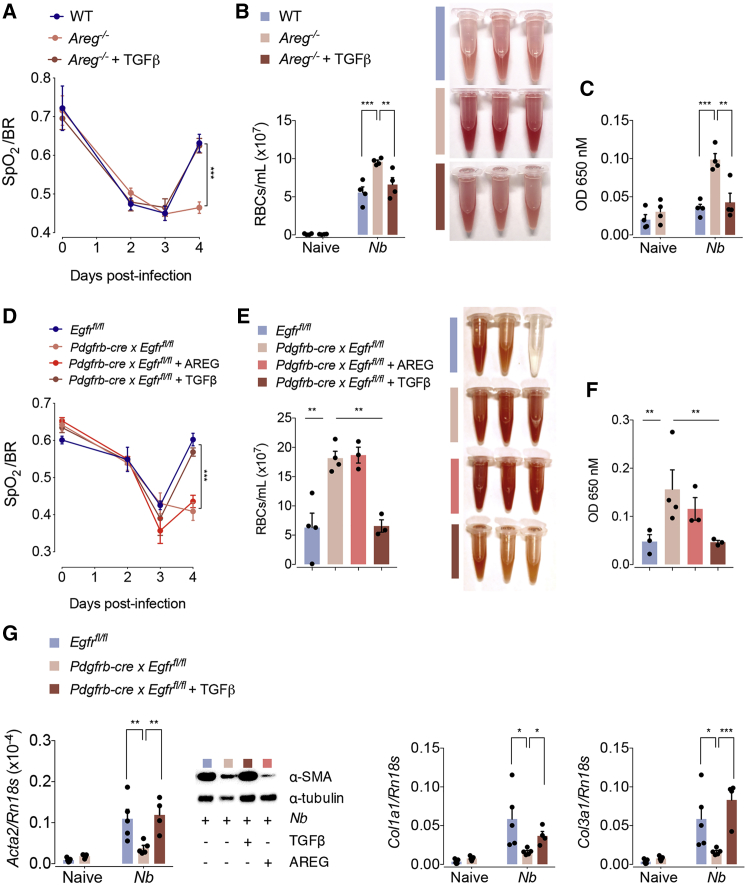
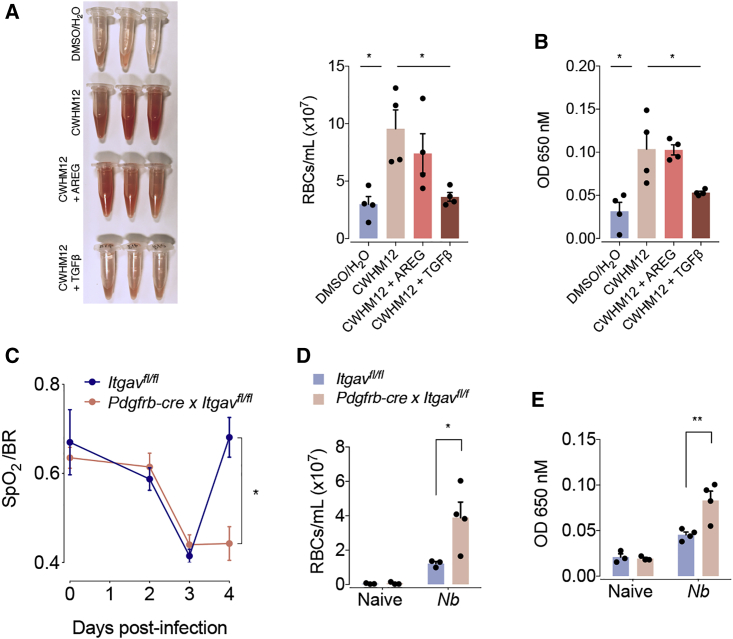
The epidermal growth factor receptor ligand Amphiregulin has a well-documented role in the restoration of tissue homeostasis after injury; however, the mechanism by which Amphiregulin contributes to wound repair remains unknown. Here we show that Amphiregulin functioned by releasing bioactive transforming growth factor beta (TGF-β) from latent complexes via integrin-αV activation. Using acute injury models in two different tissues, we found that by inducing TGF-β activation on mesenchymal stromal cells (pericytes), Amphiregulin induced their differentiation into myofibroblasts, thereby selectively contributing to the restoration of vascular barrier function within injured tissue. Furthermore, we identified macrophages as a critical source of Amphiregulin, revealing a direct effector mechanism by which these cells contribute to tissue restoration after acute injury. Combined, these observations expose a so far under-appreciated mechanism of how cells of the immune system selectively control the differentiation of tissue progenitor cells during tissue repair and inflammation.
Keywords: Amphiregulin; Macrophages; TGFb; pericytes.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Abe M., Harpel J.G., Metz C.N., Nunes I., Loskutoff D.J., Rifkin D.B. An assay for transforming growth factor-beta using cells transfected with a plasminogen activator inhibitor-1 promoter-luciferase construct. Anal. Biochem. 1994;216:276–284. - PubMed
-
- Adachi O., Kawai T., Takeda K., Matsumoto M., Tsutsui H., Sakagami M., Nakanishi K., Akira S. Targeted disruption of the MyD88 gene results in loss of IL-1- and IL-18-mediated function. Immunity. 1998;9:143–150. - PubMed
-
- Basak O., Beumer J., Wiebrands K., Seno H., van Oudenaarden A., Clevers H. Induced Quiescence of Lgr5+ Stem Cells in Intestinal Organoids Enables Differentiation of Hormone-Producing Enteroendocrine Cells. Cell Stem Cell. 2017;20:177–190.e4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
