

The Edwardsiella piscicida Type III Translocon Protein EseC Inhibits Biofilm Formation by Sequestering EseE
- PMID: 30770403
- PMCID: PMC6450016
- DOI: 10.1128/AEM.02133-18
The Edwardsiella piscicida Type III Translocon Protein EseC Inhibits Biofilm Formation by Sequestering EseE
Abstract
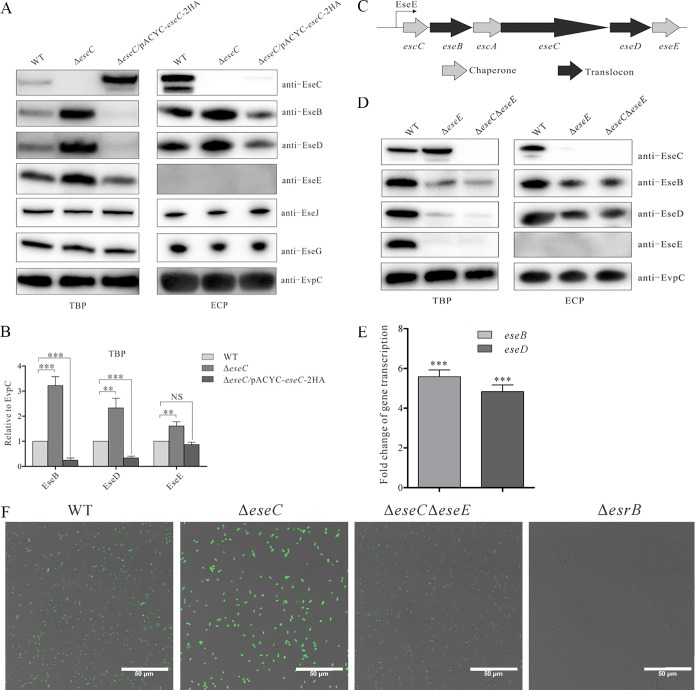
The type III secretion system (T3SS) is one of the most important virulence factors of the fish pathogen Edwardsiella piscicida It contains three translocon proteins, EseB, EseC, and EseD, required for translocation of effector proteins into host cells. We have previously shown that EseB forms filamentous appendages on the surface of E. piscicida, and these filamentous structures mediate bacterial cell-cell interactions promoting autoaggregation and biofilm formation. In the present study, we show that EseC, but not EseD, inhibits the autoaggregation and biofilm formation of E. piscicida At 18 h postsubculture, a ΔeseC strain developed strong autoaggregation and mature biofilm formation, accompanied by enhanced formation of EseB filamentous appendages. This is in contrast to the weak autoaggregation and immature biofilm formation seen in the E. piscicida wild-type strain. EseE, a protein that directly binds to EseC and also positively regulates the transcription of the escC-eseE operon, was liberated and showed increased levels in the absence of EseC. This led to augmented transcription of the escC-eseE operon, thereby increasing the steady-state protein levels of intracellular EseB, EseD, and EseE, as well as biofilm formation. Notably, the levels of intracellular EseB and EseD produced by the ΔeseE and ΔeseC ΔeseE strains were similar but remarkably lower than those produced by the wild-type strain at 18 h postsubculture. Taken together, we have shown that the translocon protein EseC inhibits biofilm formation through sequestering EseE, a positive regulator of the escC-eseE operon.IMPORTANCEEdwardsiella piscicida, previously known as Edwardsiella tarda, is a Gram-negative intracellular pathogen that mainly infects fish. The type III secretion system (T3SS) plays a pivotal role in its pathogenesis. The T3SS translocon protein EseB is required for the assembly of filamentous appendages on the surface of E. piscicida The interactions between the appendages facilitate autoaggregation and biofilm formation. In this study, we explored the role of the other two translocon proteins, EseC and EseD, in biofilm formation. We have demonstrated that EseC, but not EseD, inhibits the autoaggregation and biofilm formation of E. piscicida, providing new insights into the regulatory mechanism involved in E. piscicida biofilm formation.
Keywords: Edwardsiella piscicida; biofilm; translocon protein; type III secretion system.
Copyright © 2019 American Society for Microbiology.
Figures




References
-
- Shao S, Lai QL, Liu Q, Wu HZ, Xiao JF, Shao ZZ, Wang QY, Zhang YX. 2015. Phylogenomics characterization of a highly virulent Edwardsiella strain ET080813T encoding two distinct T3SS and three T6SS gene clusters: propose a novel species as Edwardsiella anguillarum sp. nov. Syst Appl Microbiol 38:36–47. doi: 10.1016/j.syapm.2014.10.008. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
