

Structural basis for activity of TRIC counter-ion channels in calcium release
- PMID: 30770441
- PMCID: PMC6410872
- DOI: 10.1073/pnas.1817271116
Structural basis for activity of TRIC counter-ion channels in calcium release
Abstract
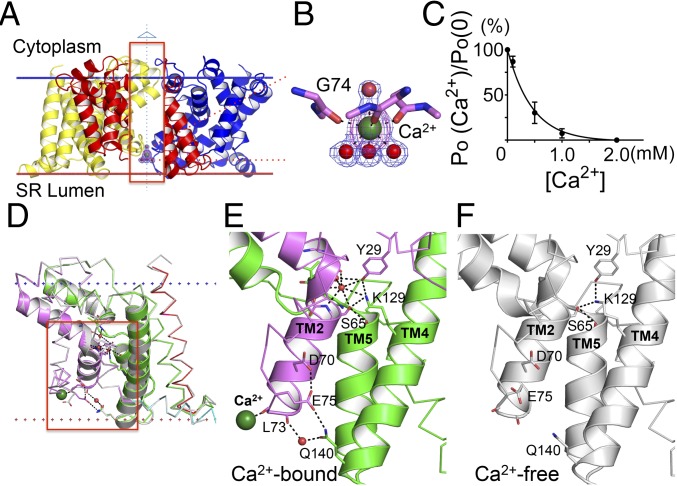
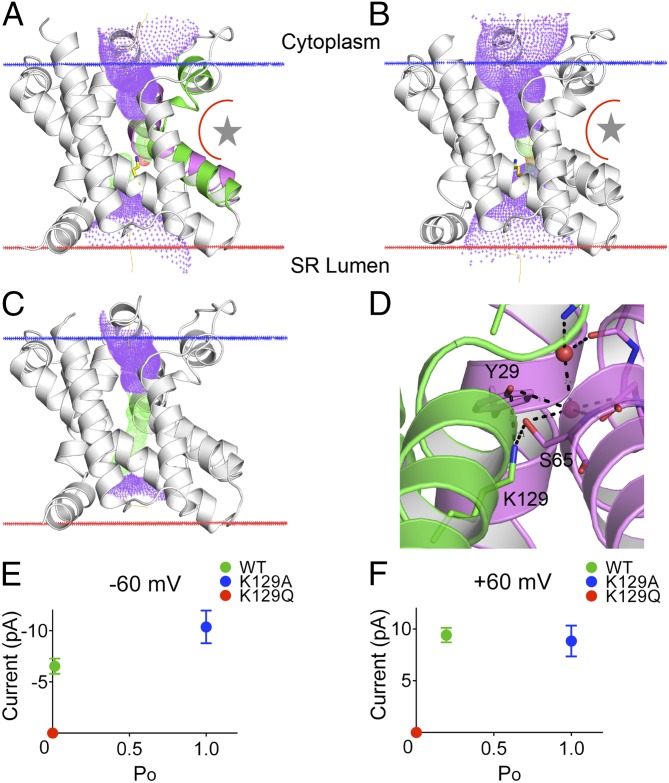
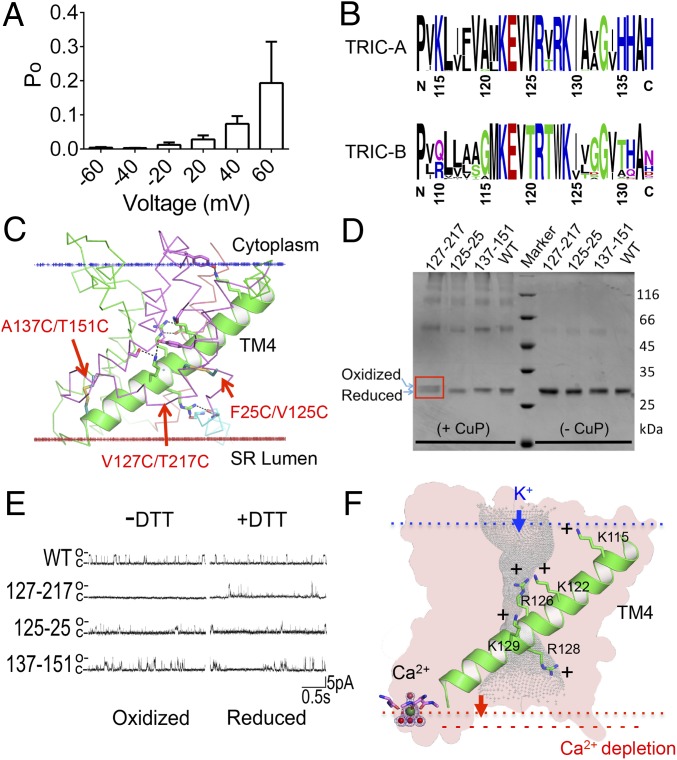
Trimeric intracellular cation (TRIC) channels are thought to provide counter-ion currents that facilitate the active release of Ca2+ from intracellular stores. TRIC activity is controlled by voltage and Ca2+ modulation, but underlying mechanisms have remained unknown. Here we describe high-resolution crystal structures of vertebrate TRIC-A and TRIC-B channels, both in Ca2+-bound and Ca2+-free states, and we analyze conductance properties in structure-inspired mutagenesis experiments. The TRIC channels are symmetric trimers, wherein we find a pore in each protomer that is gated by a highly conserved lysine residue. In the resting state, Ca2+ binding at the luminal surface of TRIC-A, on its threefold axis, stabilizes lysine blockage of the pores. During active Ca2+ release, luminal Ca2+ depletion removes inhibition to permit the lysine-bearing and voltage-sensing helix to move in response to consequent membrane hyperpolarization. Diacylglycerol is found at interprotomer interfaces, suggesting a role in metabolic control.
Keywords: Ca2+ modulation; X-ray crystallography; counter-ion mechanism; electrophysiology; lipid modulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529. - PubMed
-
- Clapham DE. Calcium signaling. Cell. 2007;131:1047–1058. - PubMed
-
- Miller C. Voltage-gated cation conductance channel from fragmented sarcoplasmic reticulum: Steady-state electrical properties. J Membr Biol. 1978;40:1–23. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
