

Stress-induced inactivation of the Staphylococcus aureus purine biosynthesis repressor leads to hypervirulence
- PMID: 30770821
- PMCID: PMC6377658
- DOI: 10.1038/s41467-019-08724-x
Stress-induced inactivation of the Staphylococcus aureus purine biosynthesis repressor leads to hypervirulence
Abstract
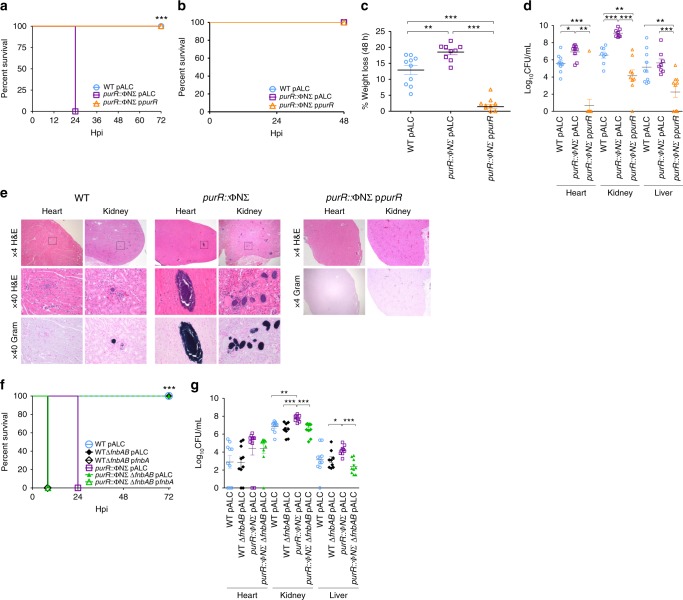
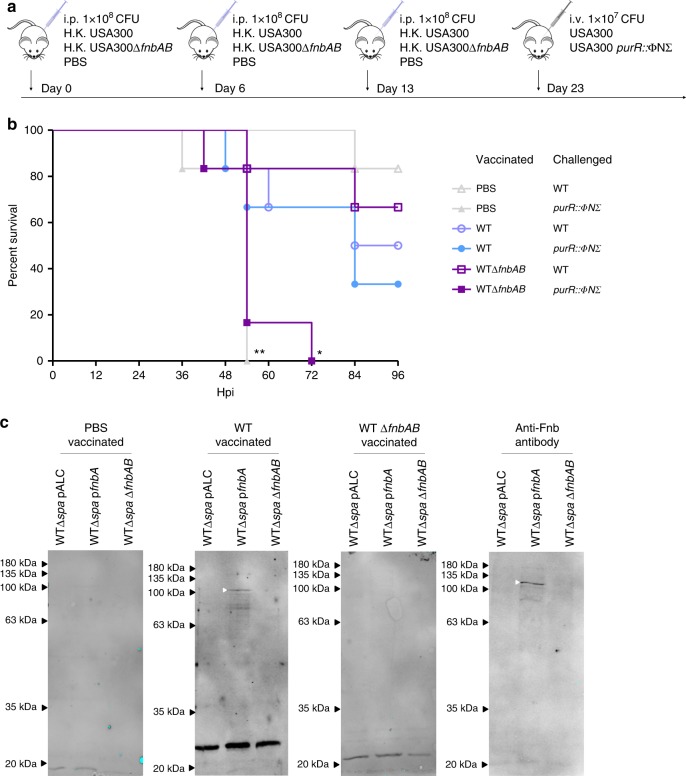
Staphylococcus aureus is a significant cause of human infection. Here, we demonstrate that mutations in the transcriptional repressor of purine biosynthesis, purR, enhance the pathogenic potential of S. aureus. Indeed, systemic infection with purR mutants causes accelerated mortality in mice, which is due to aberrant up-regulation of fibronectin binding proteins (FnBPs). Remarkably, purR mutations can arise upon exposure of S. aureus to stress, such as an intact immune system. In humans, naturally occurring anti-FnBP antibodies exist that, while not protective against recurrent S. aureus infection, ostensibly protect against hypervirulent S. aureus infections. Vaccination studies support this notion, where anti-Fnb antibodies in mice protect against purR hypervirulence. These findings provide a novel link between purine metabolism and virulence in S. aureus.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
