Loss of glutathione redox homeostasis impairs proteostasis by inhibiting autophagy-dependent protein degradation
- PMID: 30770874
- PMCID: PMC6748101
- DOI: 10.1038/s41418-018-0270-9
Loss of glutathione redox homeostasis impairs proteostasis by inhibiting autophagy-dependent protein degradation
Abstract
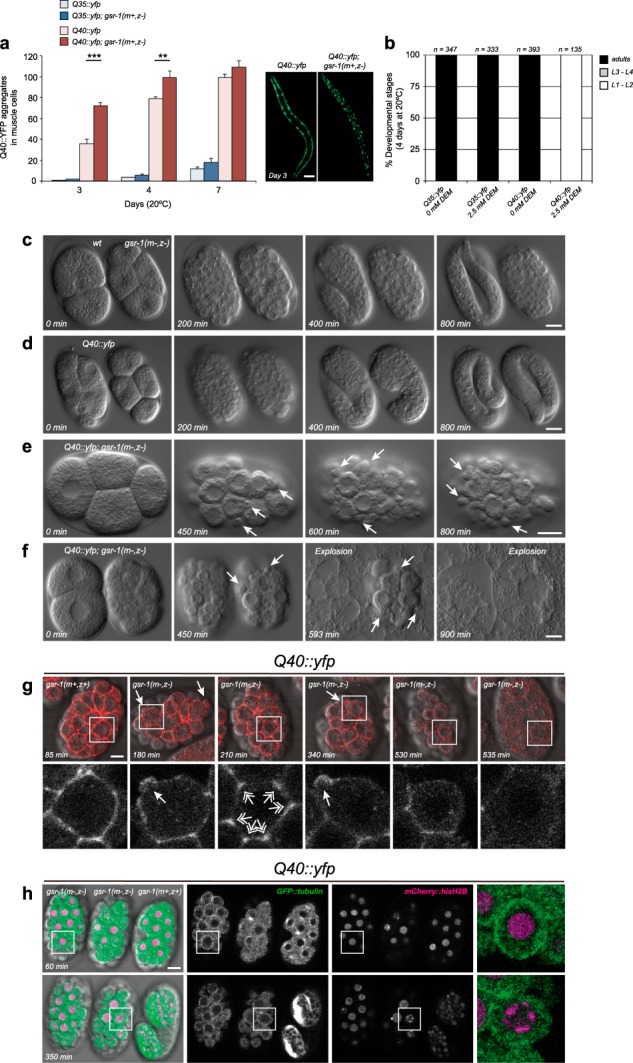
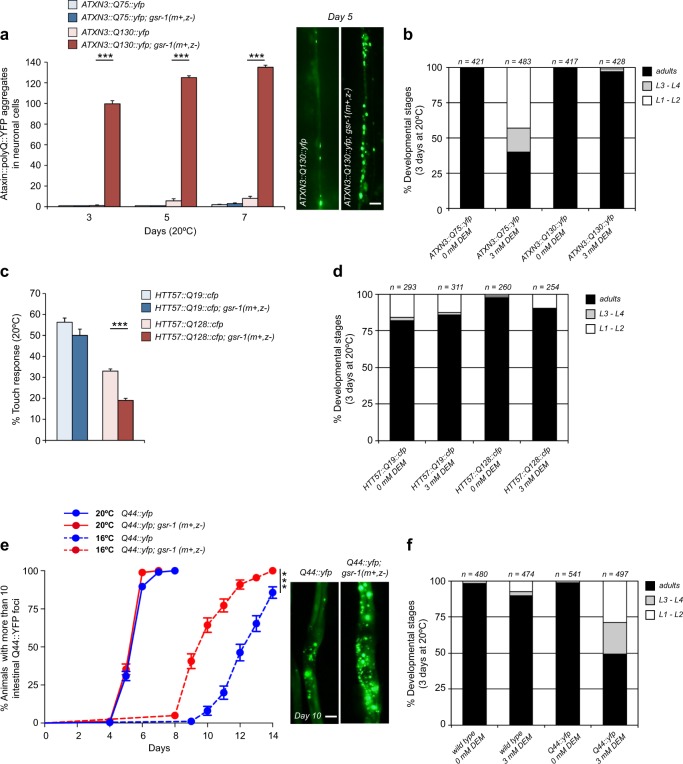
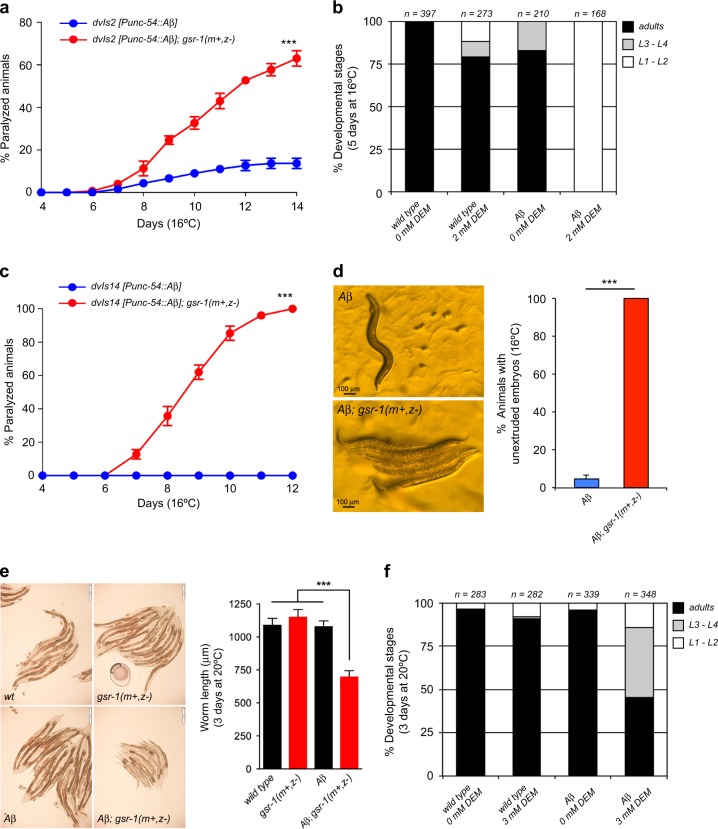
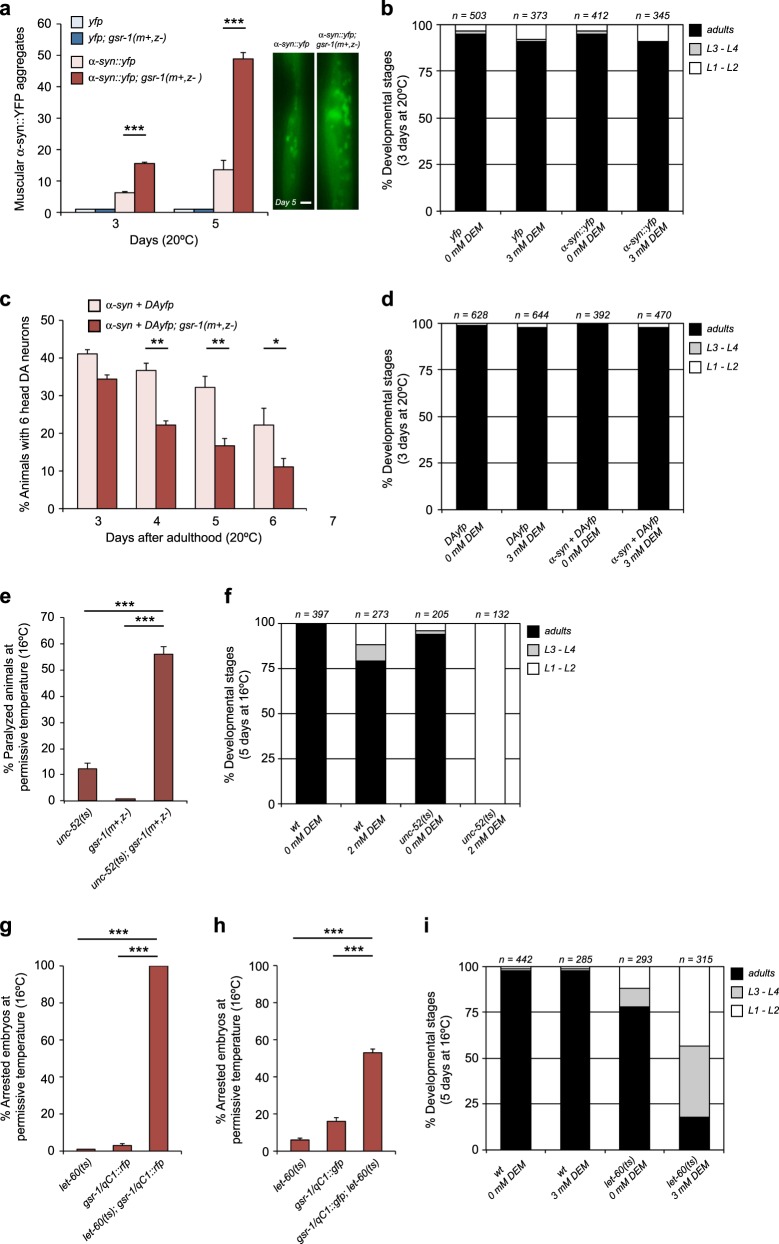
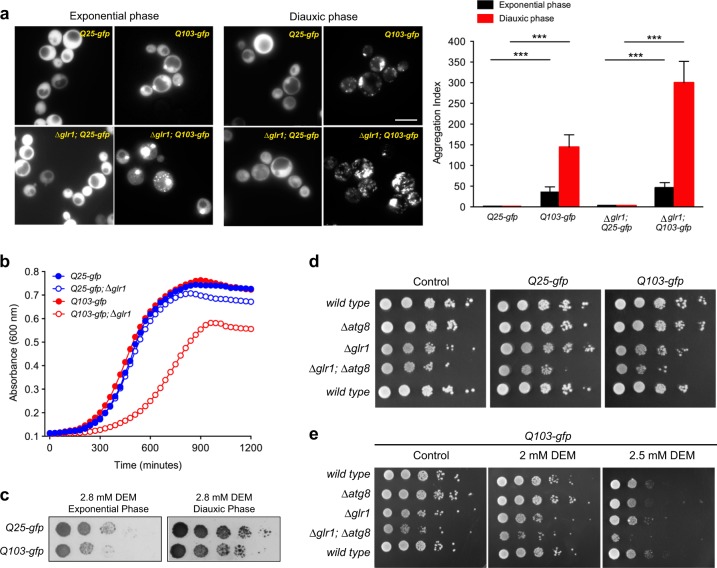
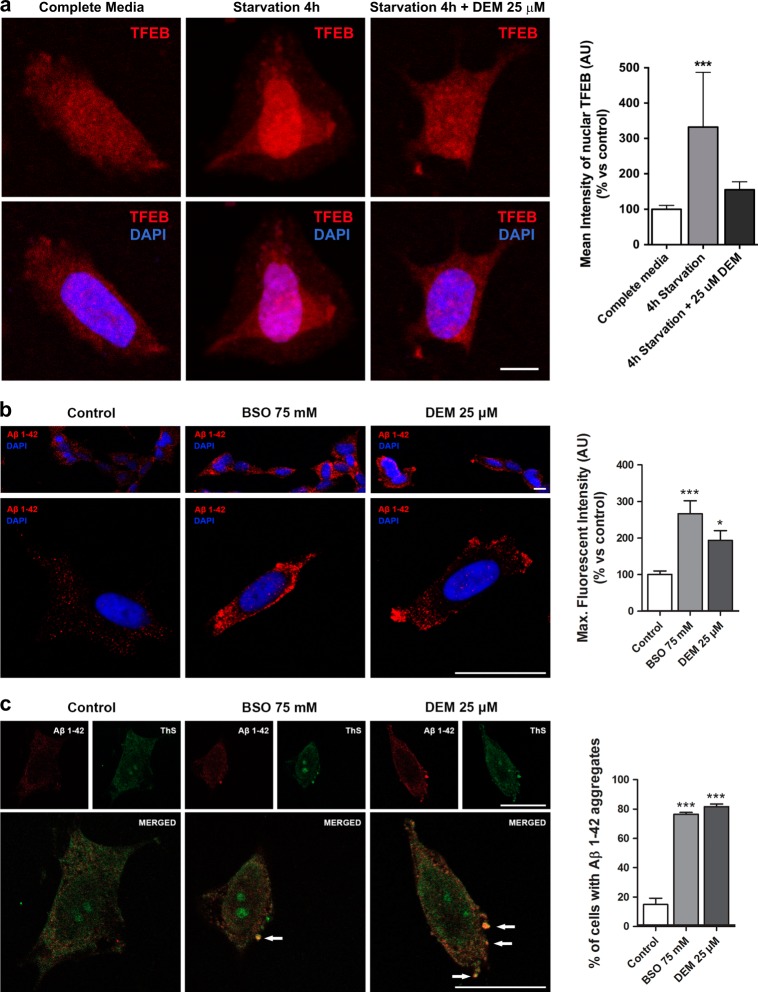
In the presence of aggregation-prone proteins, the cytosol and endoplasmic reticulum (ER) undergo a dramatic shift in their respective redox status, with the cytosol becoming more oxidized and the ER more reducing. However, whether and how changes in the cellular redox status may affect protein aggregation is unknown. Here, we show that C. elegans loss-of-function mutants for the glutathione reductase gsr-1 gene enhance the deleterious phenotypes of heterologous human, as well as endogenous worm aggregation-prone proteins. These effects are phenocopied by the GSH-depleting agent diethyl maleate. Additionally, gsr-1 mutants abolish the nuclear translocation of HLH-30/TFEB transcription factor, a key inducer of autophagy, and strongly impair the degradation of the autophagy substrate p62/SQST-1::GFP, revealing glutathione reductase may have a role in the clearance of protein aggregates by autophagy. Blocking autophagy in gsr-1 worms expressing aggregation-prone proteins results in strong synthetic developmental phenotypes and lethality, supporting the physiological importance of glutathione reductase in the regulation of misfolded protein clearance. Furthermore, impairing redox homeostasis in both yeast and mammalian cells induces toxicity phenotypes associated with protein aggregation. Together, our data reveal that glutathione redox homeostasis may be central to proteostasis maintenance through autophagy regulation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures