

Histone Acetyltransferase p300 Induces De Novo Super-Enhancers to Drive Cellular Senescence
- PMID: 30773298
- PMCID: PMC6688479
- DOI: 10.1016/j.molcel.2019.01.021
Histone Acetyltransferase p300 Induces De Novo Super-Enhancers to Drive Cellular Senescence
Abstract
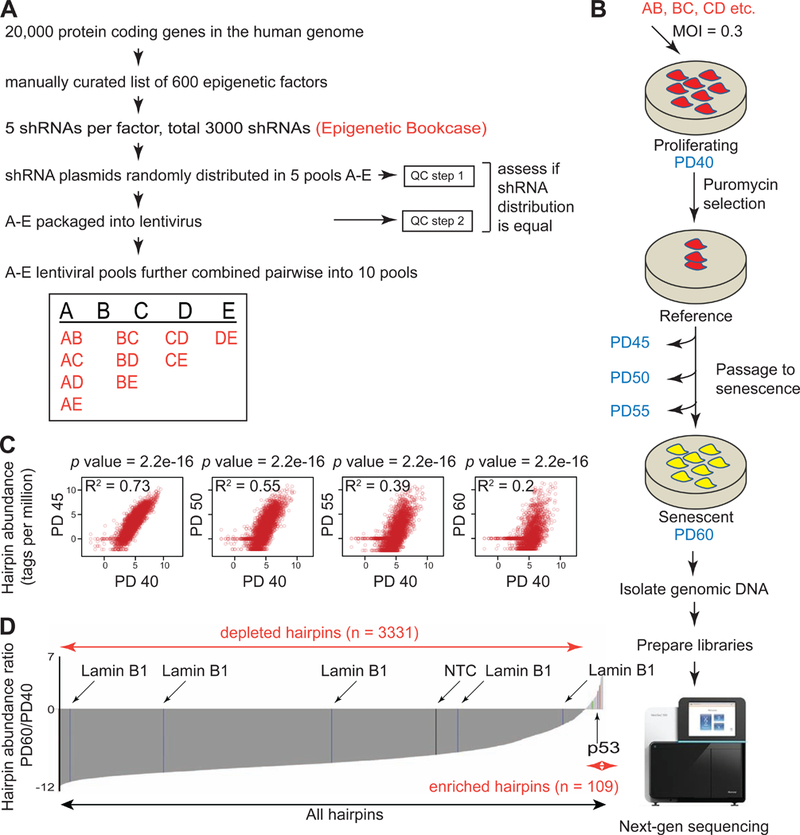
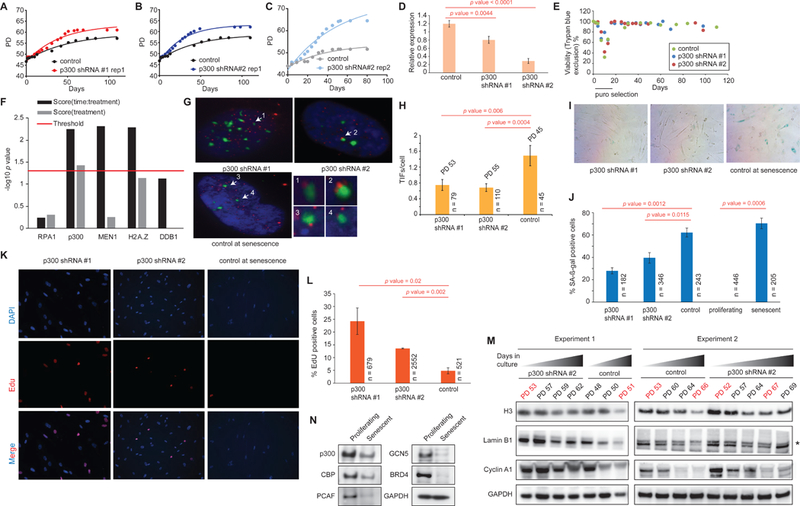
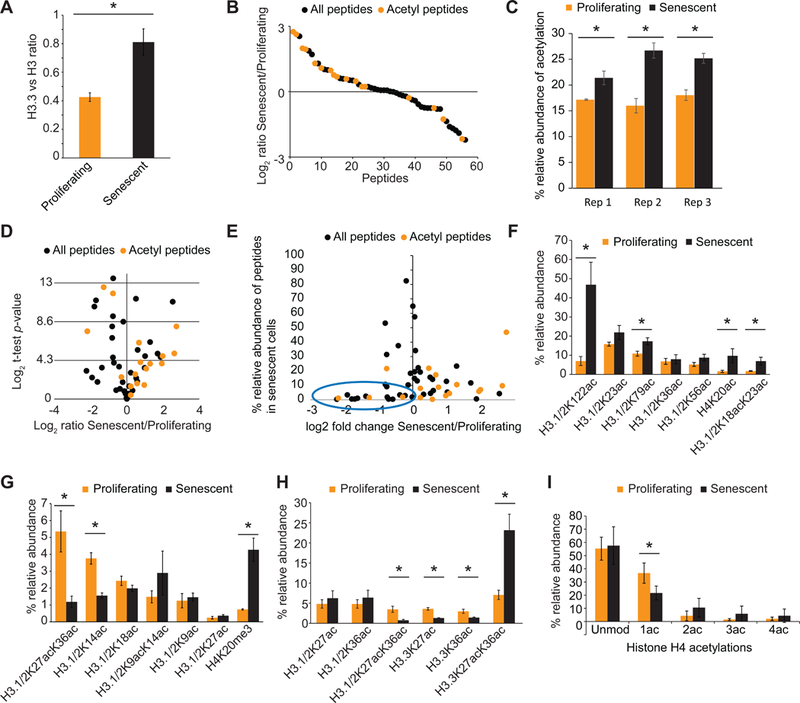
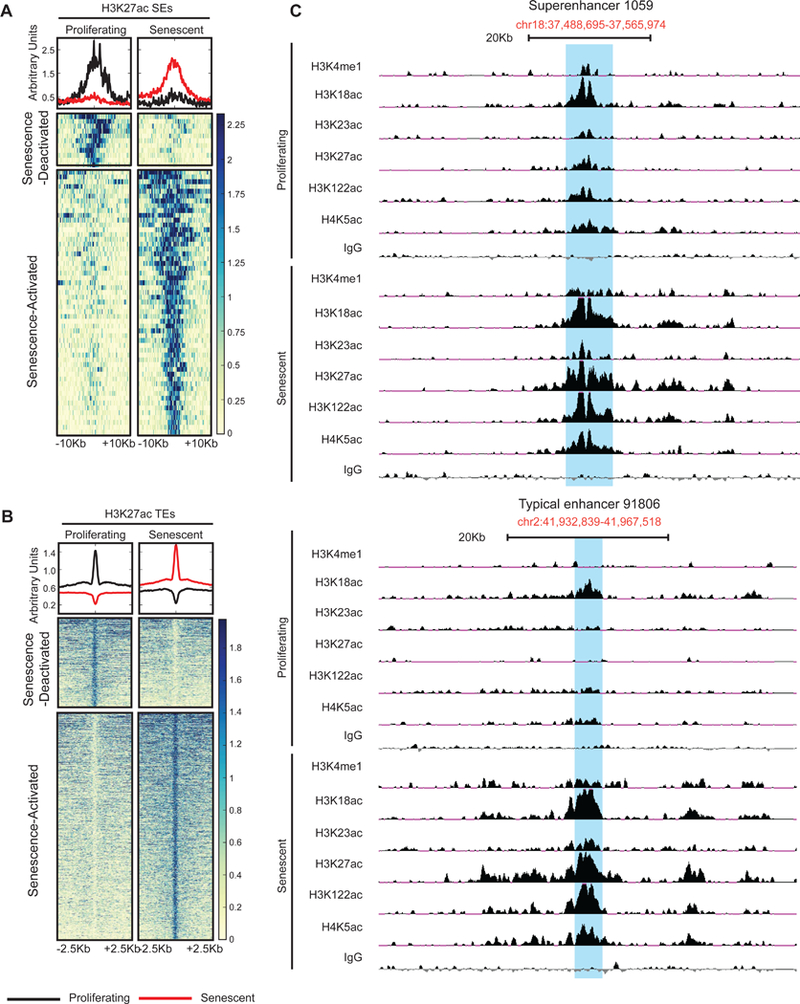
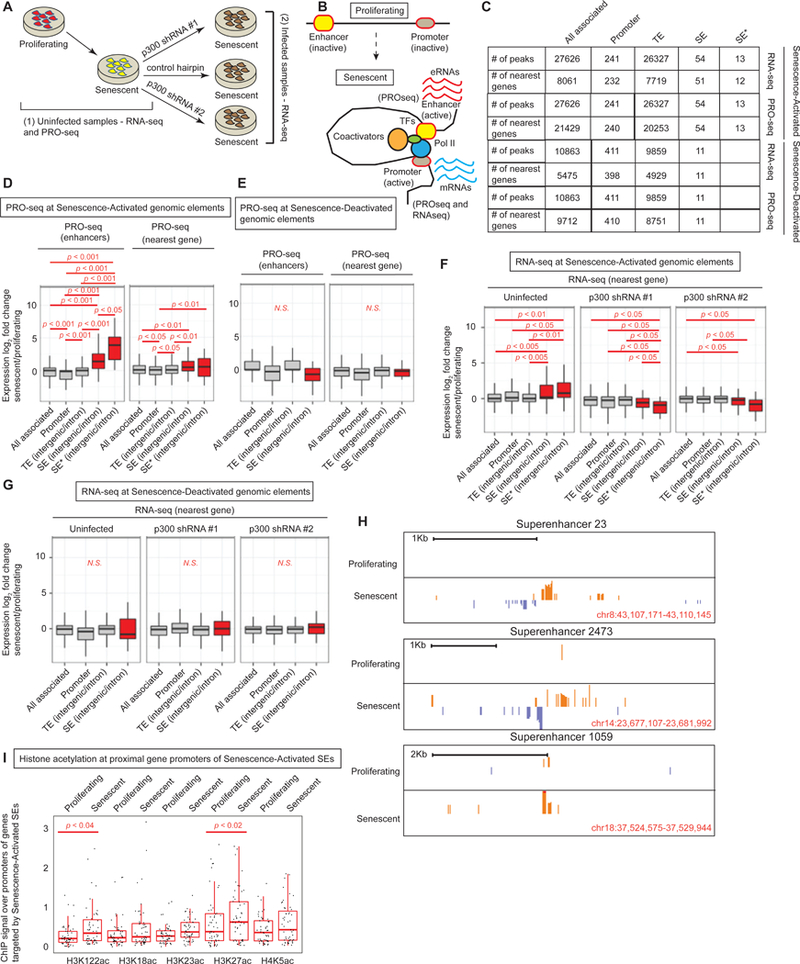
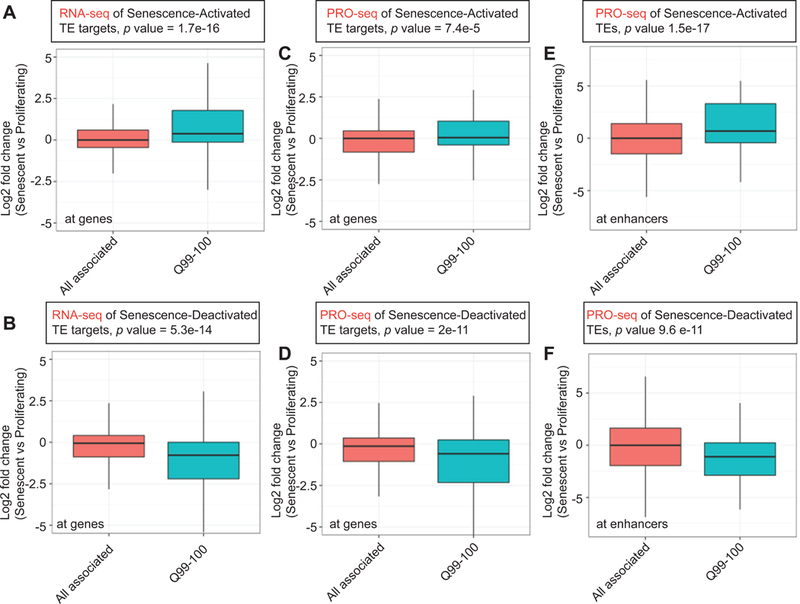
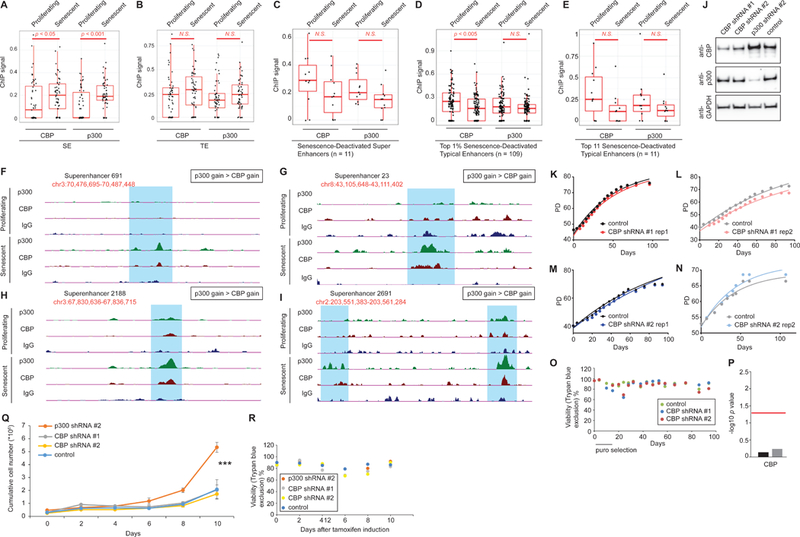
Accumulation of senescent cells during aging contributes to chronic inflammation and age-related diseases. While senescence is associated with profound alterations of the epigenome, a systematic view of epigenetic factors in regulating senescence is lacking. Here, we curated a library of short hairpin RNAs for targeted silencing of all known epigenetic proteins and performed a high-throughput screen to identify key candidates whose downregulation can delay replicative senescence of primary human cells. This screen identified multiple new players including the histone acetyltransferase p300 that was found to be a primary driver of the senescent phenotype. p300, but not the paralogous CBP, induces a dynamic hyper-acetylated chromatin state and promotes the formation of active enhancer elements in the non-coding genome, leading to a senescence-specific gene expression program. Our work illustrates a causal role of histone acetyltransferases and acetylation in senescence and suggests p300 as a potential therapeutic target for senescence and age-related diseases.
Keywords: chromatin; enhancers; epigenetics; p300; senescence.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
Comment in
-
The unusual SASPects.Nat Rev Mol Cell Biol. 2019 Apr;20(4):195. doi: 10.1038/s41580-019-0111-9. Nat Rev Mol Cell Biol. 2019. PMID: 30770903 No abstract available.
References
-
- Akaike H (1974). A new look at the statistical model identification. IEEE Transactions on Automatic Control 19, 716–723.
-
- Atlasi Y, and Stunnenberg HG (2017). The interplay of epigenetic marks during stem cell differentiation and development. Nat Rev Genet. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
