

Detection of Low-Abundance Metabolites in Live Cells Using an RNA Integrator
- PMID: 30773480
- PMCID: PMC6474789
- DOI: 10.1016/j.chembiol.2019.01.005
Detection of Low-Abundance Metabolites in Live Cells Using an RNA Integrator
Abstract
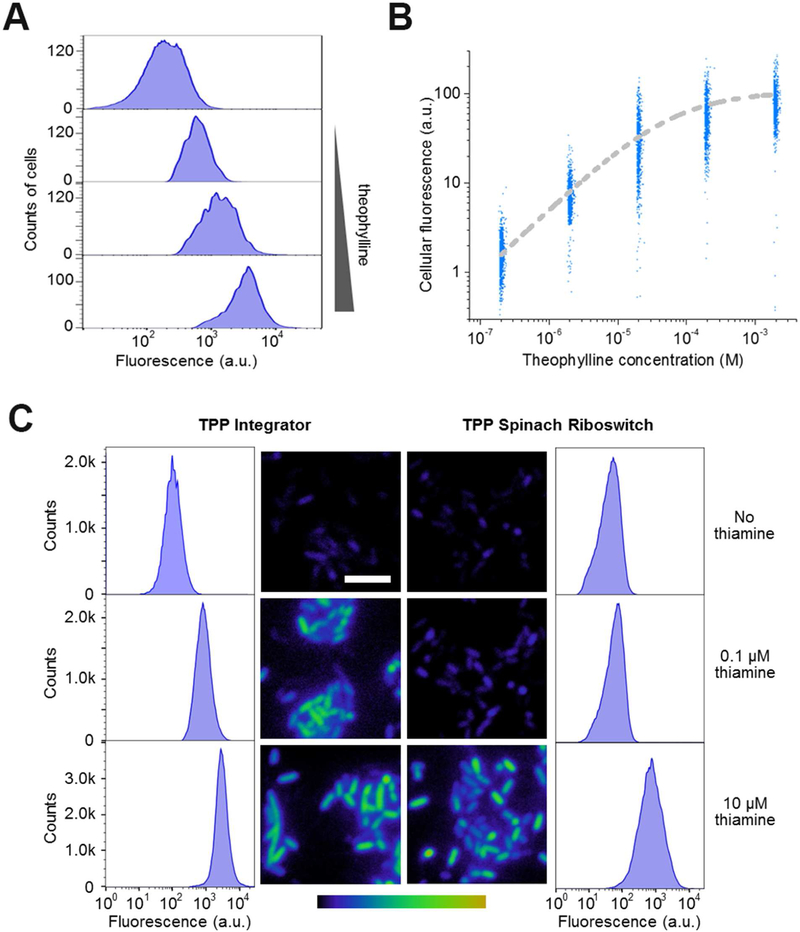
Genetically encoded biosensors are useful tools for detecting the presence and levels of diverse biomolecules in living cells. However, low-abundance targets are difficult to detect because they are often unable to bind and activate enough biosensors to detect using standard microscopic imaging approaches. Here we describe a type of RNA-based biosensor, an RNA integrator, which enables detection of low-abundance targets in vitro and in living cells. The RNA integrator is an RNA sequence comprising a ribozyme and an unfolded form of the fluorogenic aptamer Broccoli. Upon binding its target, the ribozyme undergoes cleavage and releases Broccoli, which subsequently folds and becomes fluorescent. Importantly, each target molecule can bind and induce cleavage of multiple copies of the integrator sensor, resulting in an amplified signal. We show that this approach can be generalized to numerous different ribozyme types for the detection of various small molecules.
Keywords: RNA probes; cellular imaging; fluorescence; fluorogenic aptamer; low-abundance metabolite; ribozyme.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
S.R.J. is a co-founder of Lucerna Technologies and has equity in this company. Lucerna has licensed commercialization of technology related to Spinach and other RNA–fluorophore complexes.
Figures
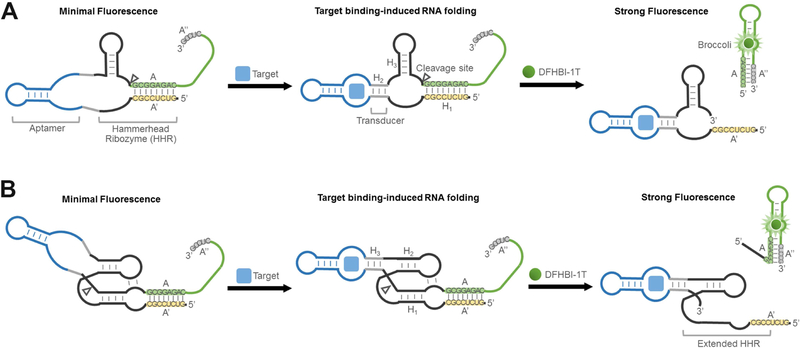
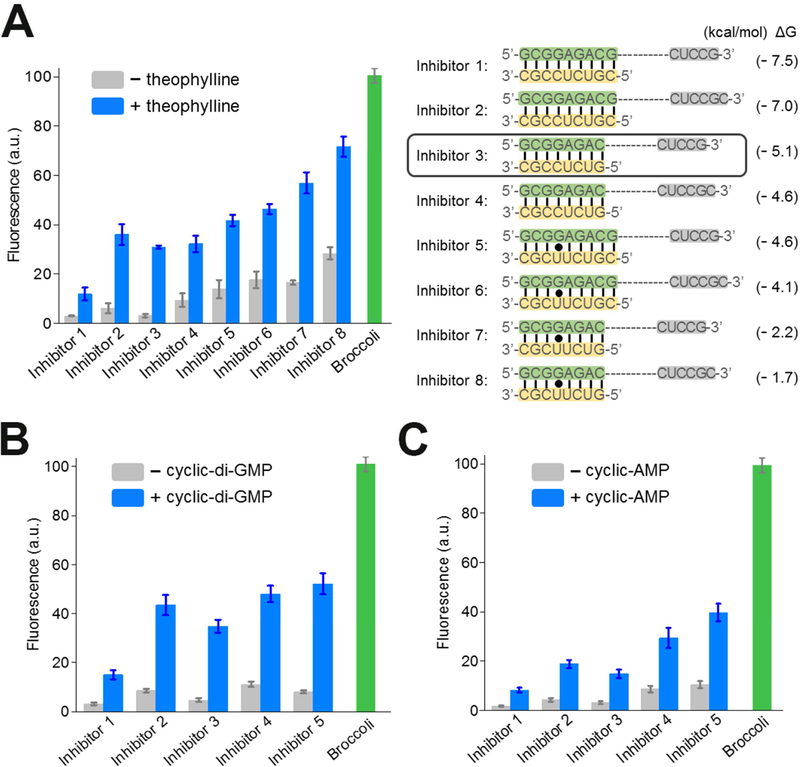
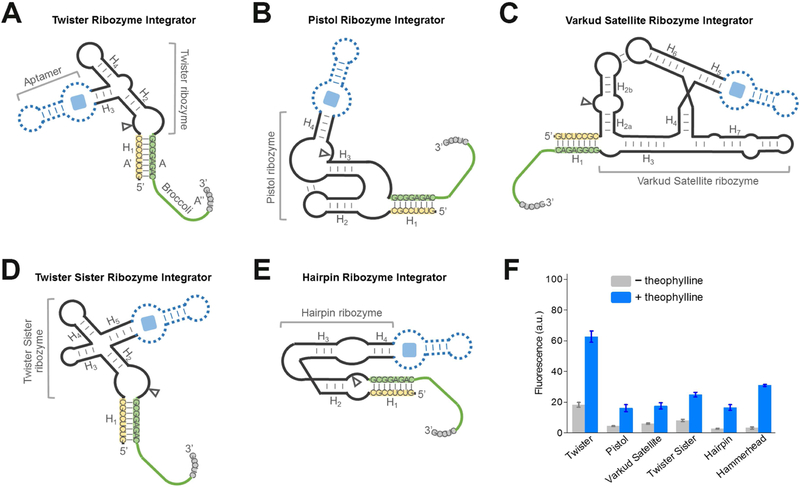
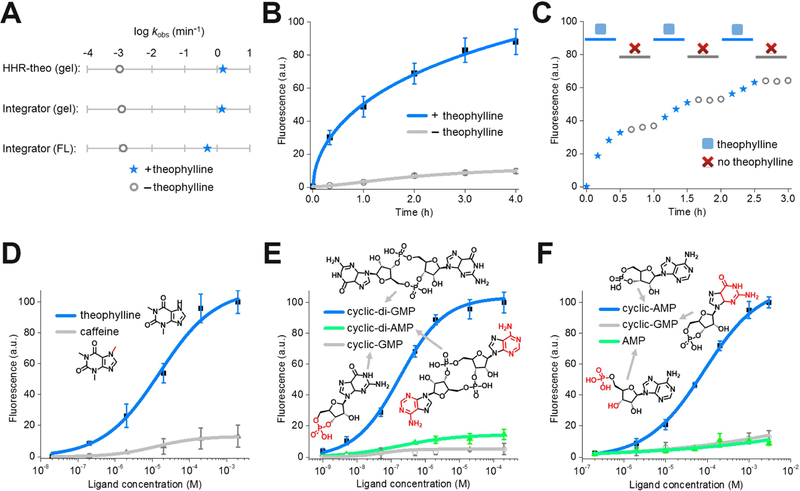
Comment in
-
Adding Broccoli to the Biosensor Menu.Cell Chem Biol. 2019 Apr 18;26(4):463-465. doi: 10.1016/j.chembiol.2019.04.004. Cell Chem Biol. 2019. PMID: 31002799
References
-
- Beilstein K, Wittmann A, Grez M, and Suess B (2015). Conditional control of mammalian gene expression by tetracycline-dependent hammerhead ribozymes. ACS Synth. Biol 4, 526–534. - PubMed
-
- Birikh KR, Heaton PA, and Eckstein F (1997). The structure, function and application of the hammerhead ribozyme. Eur. J. Biochem 245, 1–16. - PubMed
-
- Breaker RR (2002). Engineered allosteric ribozymes as a biosensor components. Curr. Opin. Biotechnol 13, 31–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
