

Eye Movement Compensation and Spatial Updating in Visual Prosthetics: Mechanisms, Limitations and Future Directions
- PMID: 30774585
- PMCID: PMC6368147
- DOI: 10.3389/fnsys.2018.00073
Eye Movement Compensation and Spatial Updating in Visual Prosthetics: Mechanisms, Limitations and Future Directions
Abstract
Despite appearing automatic and effortless, perceiving the visual world is a highly complex process that depends on intact visual and oculomotor function. Understanding the mechanisms underlying spatial updating (i.e., gaze contingency) represents an important, yet unresolved issue in the fields of visual perception and cognitive neuroscience. Many questions regarding the processes involved in updating visual information as a function of the movements of the eyes are still open for research. Beyond its importance for basic research, gaze contingency represents a challenge for visual prosthetics as well. While most artificial vision studies acknowledge its importance in providing accurate visual percepts to the blind implanted patients, the majority of the current devices do not compensate for gaze position. To-date, artificial percepts to the blind population have been provided either by intraocular light-sensing circuitry or by using external cameras. While the former commonly accounts for gaze shifts, the latter requires the use of eye-tracking or similar technology in order to deliver percepts based on gaze position. Inspired by the need to overcome the hurdle of gaze contingency in artificial vision, we aim to provide a thorough overview of the research addressing the neural underpinnings of eye compensation, as well as its relevance in visual prosthetics. The present review outlines what is currently known about the mechanisms underlying spatial updating and reviews the attempts of current visual prosthetic devices to overcome the hurdle of gaze contingency. We discuss the limitations of the current devices and highlight the need to use eye-tracking methodology in order to introduce gaze-contingent information to visual prosthetics.
Keywords: artificial vision; blindness; eye movements; gaze contingency; neuroprosthetics; visual prosthetics.
Figures
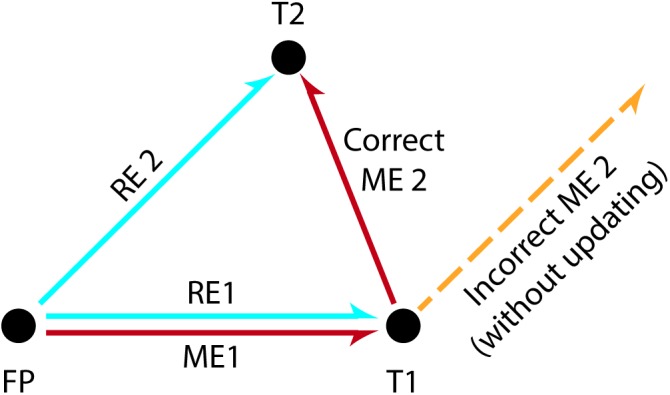
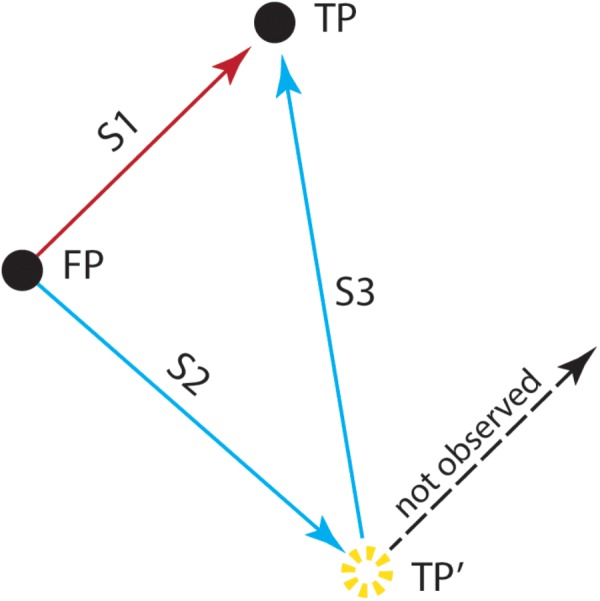
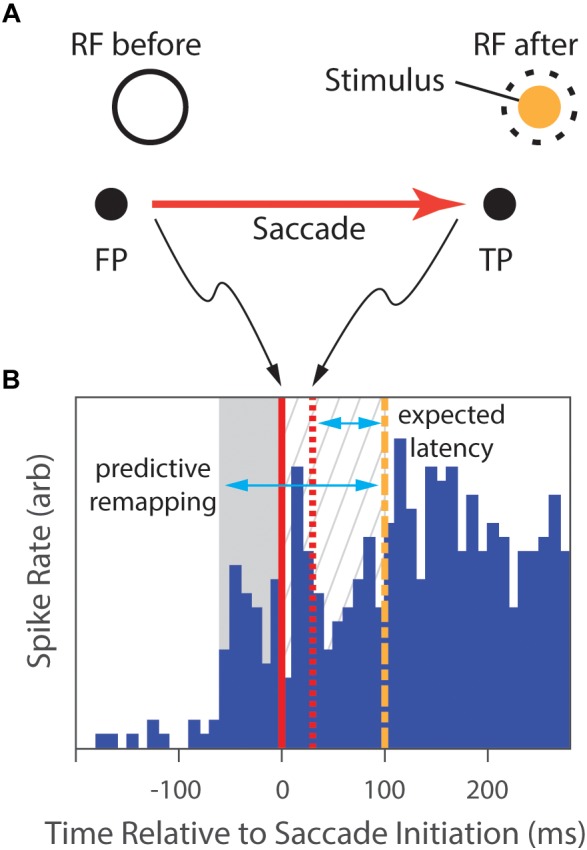
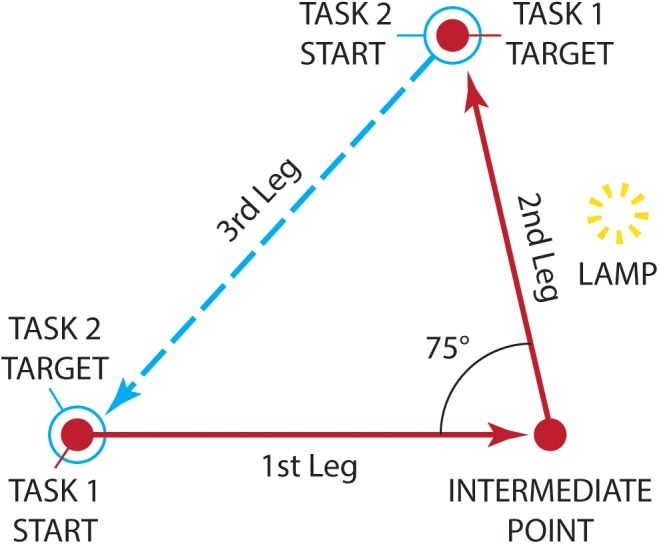
References
-
- Andersen R. A., Gnadt J. W. (1989). “Posterior parietal cortex,” in The Neurobiology of Saccadic Eye Movements, Reviews of Oculomotor Research, Vol. III, eds Wurtz R. H., Goldberg M. E. (Amsterdam: Elsevier; ), 315–336.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources