

Metabolism as an early predictor of DPSCs aging
- PMID: 30778087
- PMCID: PMC6379364
- DOI: 10.1038/s41598-018-37489-4
Metabolism as an early predictor of DPSCs aging
Abstract
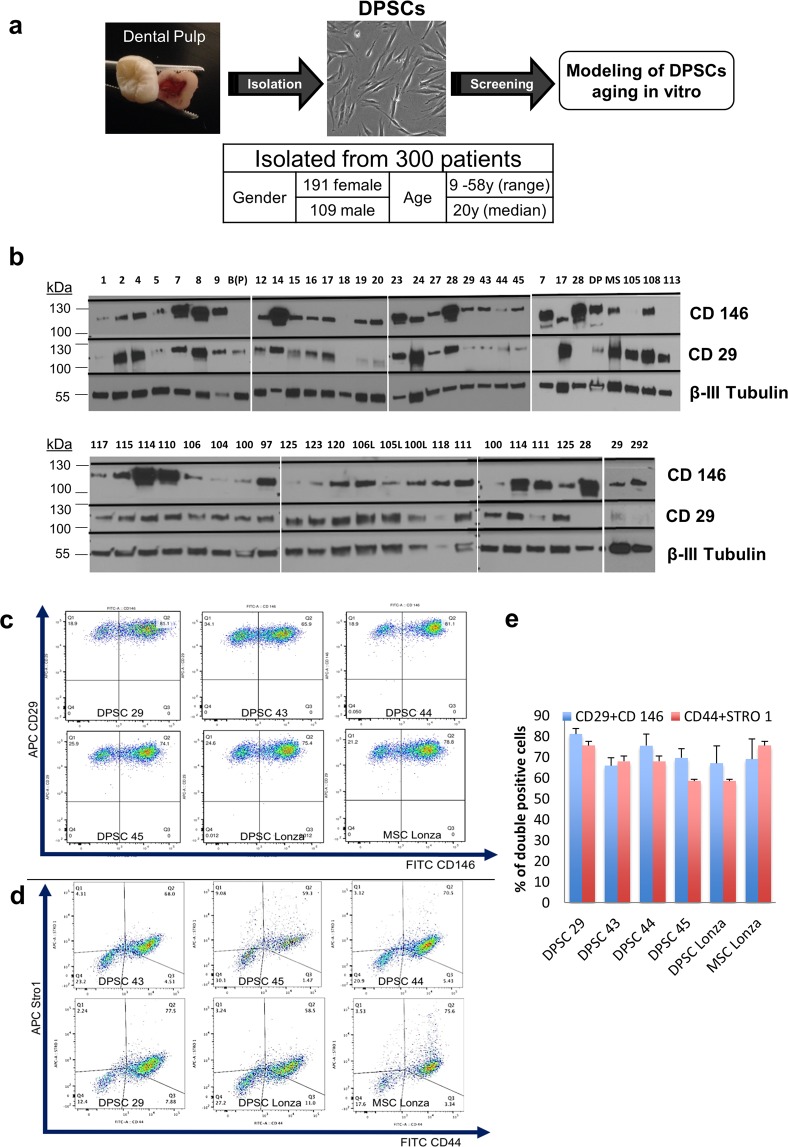
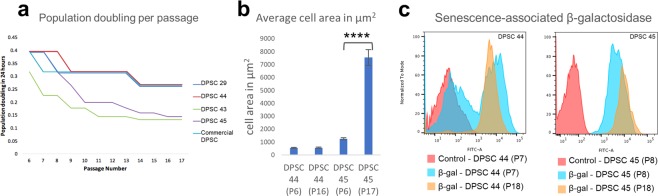
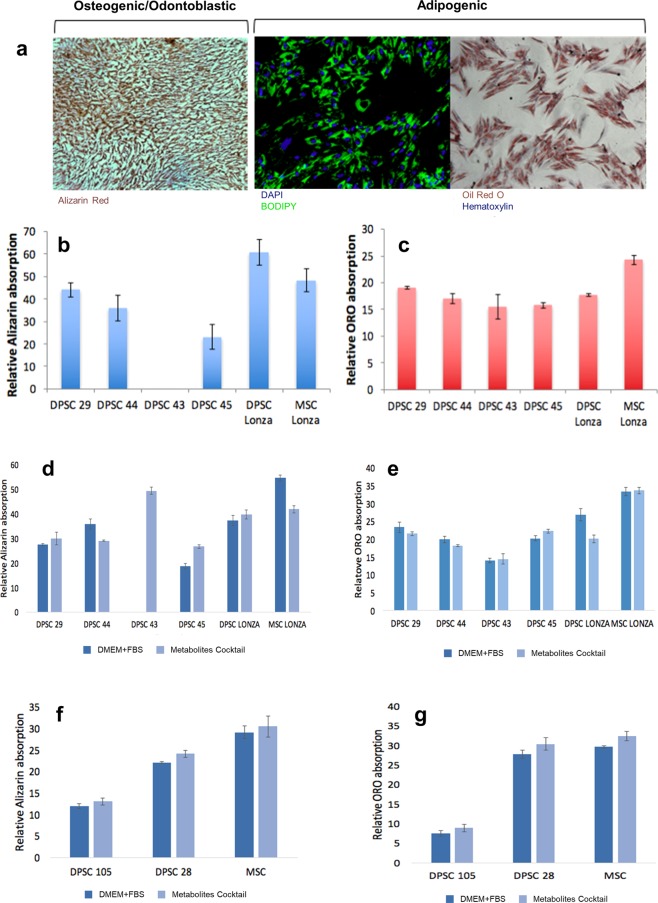
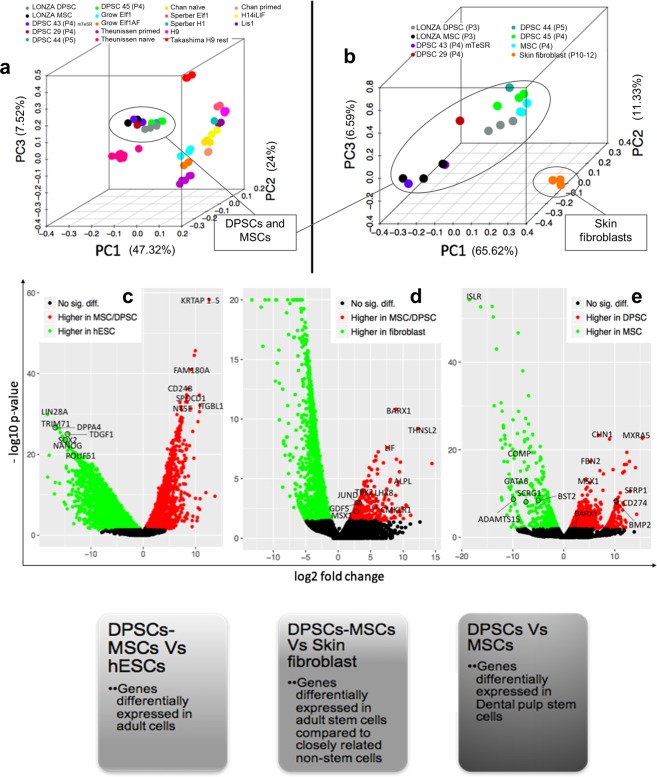
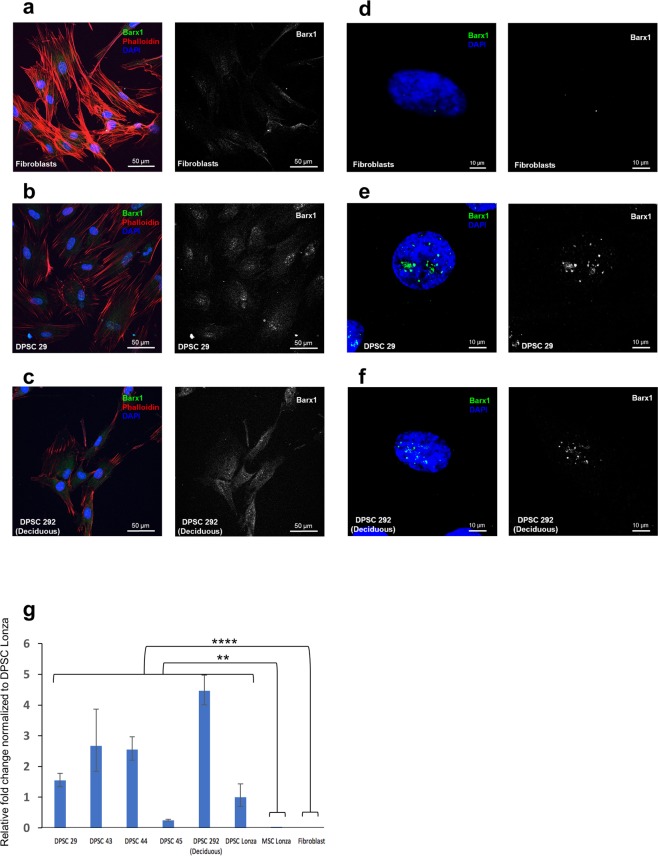
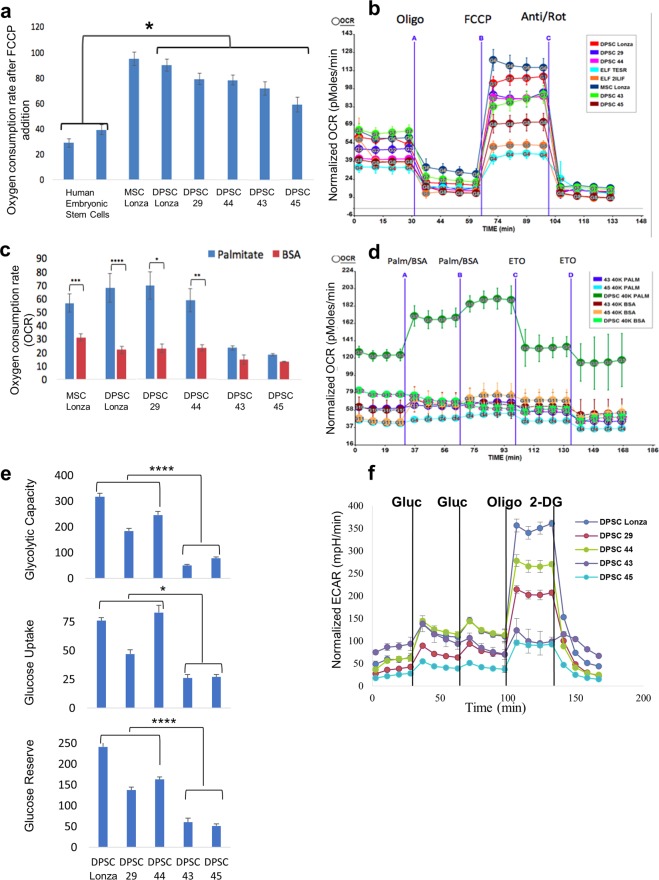
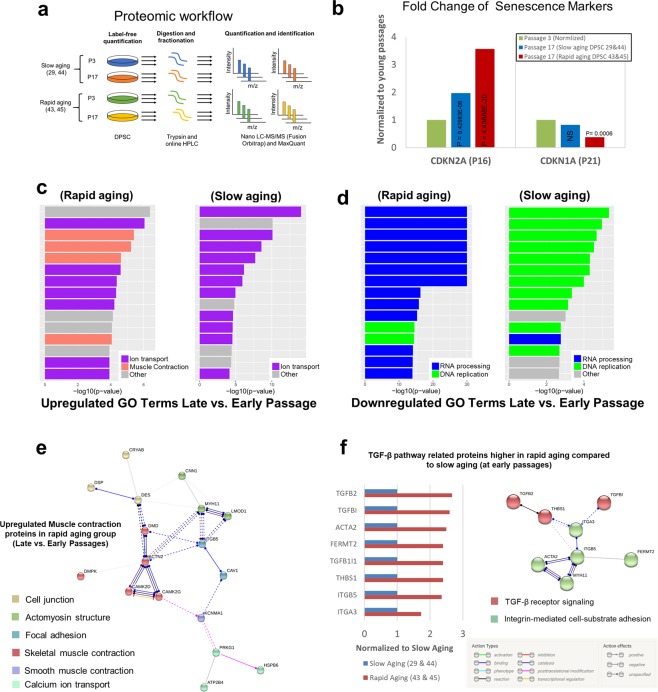
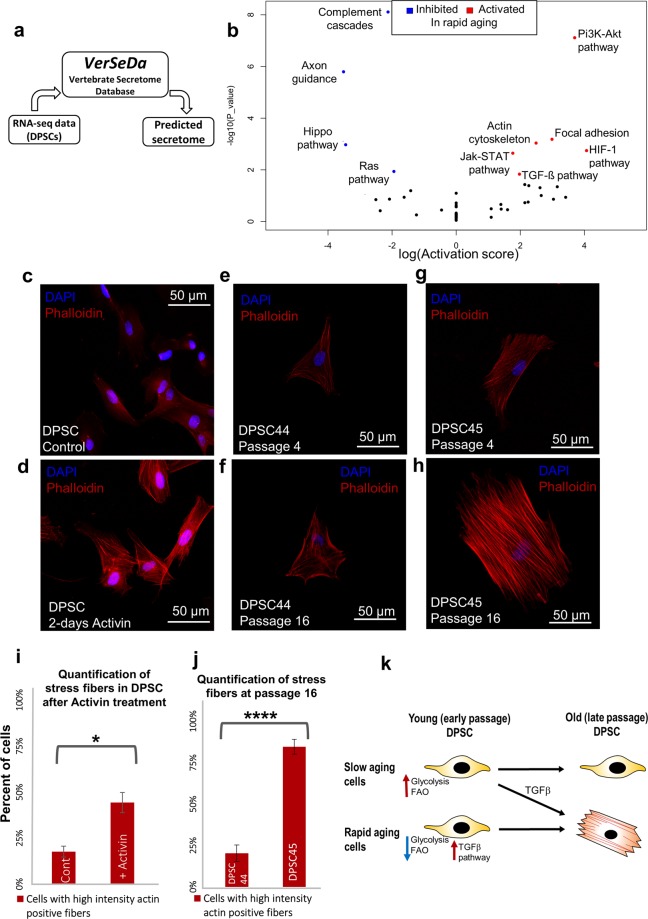
Tissue resident adult stem cells are known to participate in tissue regeneration and repair that follows cell turnover, or injury. It has been well established that aging impedes the regeneration capabilities at the cellular level, but it is not clear if the different onset of stem cell aging between individuals can be predicted or prevented at an earlier stage. Here we studied the dental pulp stem cells (DPSCs), a population of adult stem cells that is known to participate in the repair of an injured tooth, and its properties can be affected by aging. The dental pulp from third molars of a diverse patient group were surgically extracted, generating cells that had a high percentage of mesenchymal stem cell markers CD29, CD44, CD146 and Stro1 and had the ability to differentiate into osteo/odontogenic and adipogenic lineages. Through RNA seq and qPCR analysis we identified homeobox protein, Barx1, as a marker for DPSCs. Furthermore, using high throughput transcriptomic and proteomic analysis we identified markers for DPSC populations with accelerated replicative senescence. In particular, we show that the transforming growth factor-beta (TGF-β) pathway and the cytoskeletal proteins are upregulated in rapid aging DPSCs, indicating a loss of stem cell characteristics and spontaneous initiation of terminal differentiation. Importantly, using metabolic flux analysis, we identified a metabolic signature for the rapid aging DPSCs, prior to manifestation of senescence phenotypes. This metabolic signature therefore can be used to predict the onset of replicative senescence. Hence, the present study identifies Barx1 as a DPSCs marker and dissects the first predictive metabolic signature for DPSCs aging.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Analysis of Senescence-Related Differentiation Potentials and Gene Expression Profiles in Human Dental Pulp Stem Cells.Cells Tissues Organs. 2017;203(1):1-11. doi: 10.1159/000448026. Epub 2016 Sep 15. Cells Tissues Organs. 2017. PMID: 27627434
-
FAM96B inhibits the senescence of dental pulp stem cells.Cell Biol Int. 2020 May;44(5):1193-1203. doi: 10.1002/cbin.11319. Epub 2020 Feb 19. Cell Biol Int. 2020. PMID: 32039527
-
Comparative proteomic profiling of human dental pulp stem cells and periodontal ligament stem cells under in vitro osteogenic induction.Arch Oral Biol. 2018 May;89:9-19. doi: 10.1016/j.archoralbio.2018.01.015. Epub 2018 Feb 3. Arch Oral Biol. 2018. PMID: 29407636
-
Expression of Odontogenic and Osteogenic Markers in DPSCs and SHED: A Review.Curr Stem Cell Res Ther. 2017;12(1):71-79. doi: 10.2174/1574888x11666160815095733. Curr Stem Cell Res Ther. 2017. PMID: 27527527 Review.
-
Cellular senescence in dental pulp stem cells.Arch Oral Biol. 2019 Mar;99:150-155. doi: 10.1016/j.archoralbio.2019.01.012. Epub 2019 Jan 17. Arch Oral Biol. 2019. PMID: 30685471 Review.
Cited by
-
Single-cell census of human tooth development enables generation of human enamel.Dev Cell. 2023 Oct 23;58(20):2163-2180.e9. doi: 10.1016/j.devcel.2023.07.013. Epub 2023 Aug 14. Dev Cell. 2023. PMID: 37582367 Free PMC article.
-
Epigenetic Regulation of Dental Pulp Stem Cell Fate.Stem Cells Int. 2020 Oct 13;2020:8876265. doi: 10.1155/2020/8876265. eCollection 2020. Stem Cells Int. 2020. PMID: 33149742 Free PMC article. Review.
-
Public RNA-seq data-based identification and functional analyses reveal that MXRA5 retains proliferative and migratory abilities of dental pulp stem cells.Sci Rep. 2023 Sep 20;13(1):15574. doi: 10.1038/s41598-023-42684-z. Sci Rep. 2023. PMID: 37730838 Free PMC article.
-
Metabolic shift and the effect of mitochondrial respiration on the osteogenic differentiation of dental pulp stem cells.PeerJ. 2023 Apr 21;11:e15164. doi: 10.7717/peerj.15164. eCollection 2023. PeerJ. 2023. PMID: 37101792 Free PMC article.
-
Downregulation of ROR2 promotes dental pulp stem cell senescence by inhibiting STK4-FOXO1/SMS1 axis in sphingomyelin biosynthesis.Aging Cell. 2021 Aug;20(8):e13430. doi: 10.1111/acel.13430. Epub 2021 Jul 18. Aging Cell. 2021. PMID: 34278704 Free PMC article.
References
-
- Soteriou, D. & Fuchs, Y. A matter of life and death: stem cell survival in tissue regeneration and tumour formation. Nature reviews. Cancer, 10.1038/nrc.2017.122 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
