

NAD+ metabolism governs the proinflammatory senescence-associated secretome
- PMID: 30778219
- PMCID: PMC6448588
- DOI: 10.1038/s41556-019-0287-4
NAD+ metabolism governs the proinflammatory senescence-associated secretome
Abstract
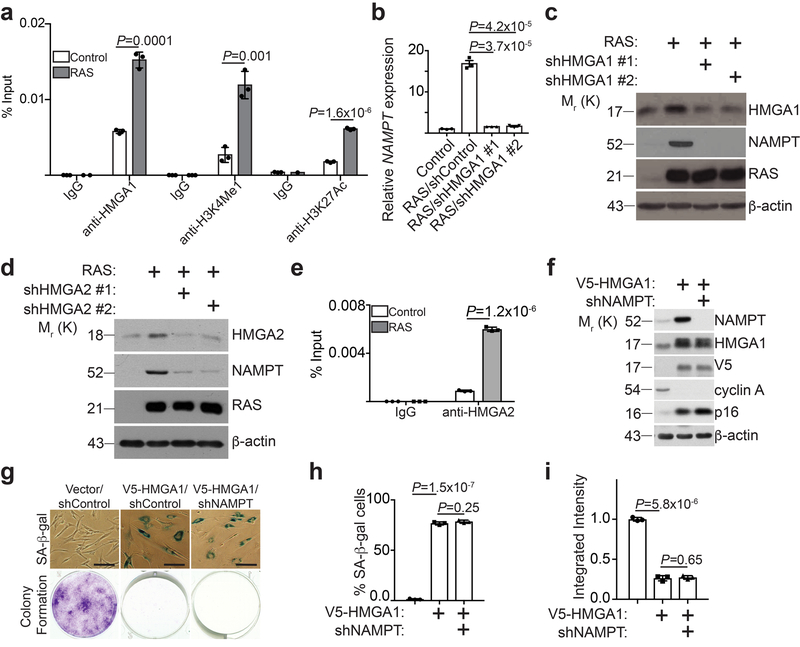
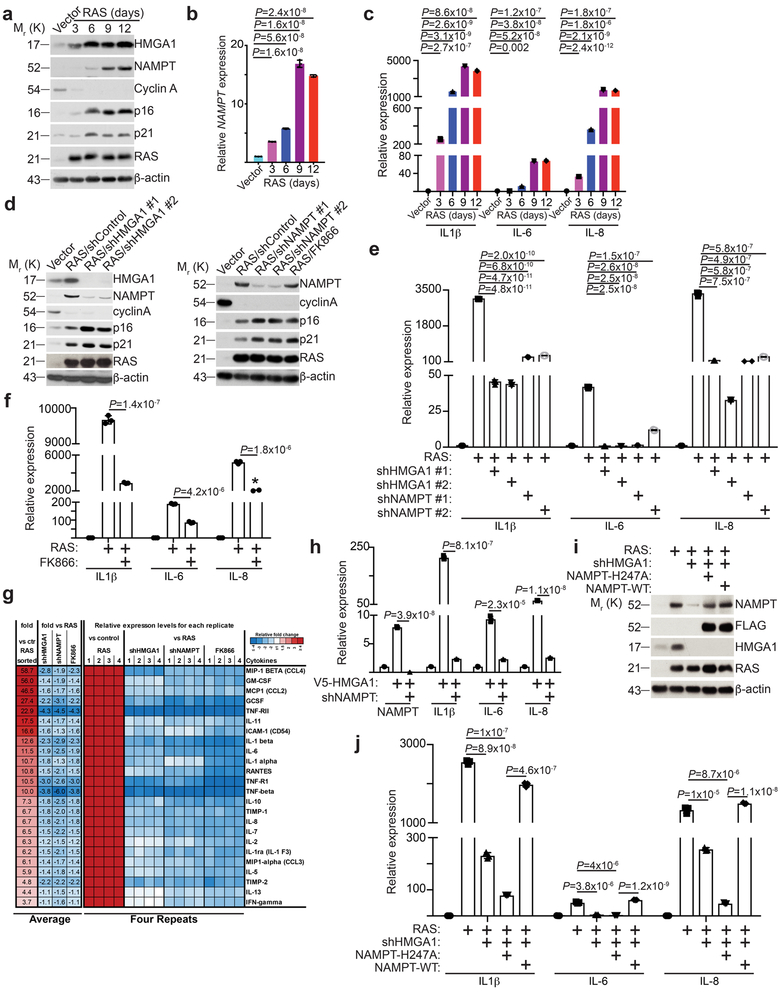
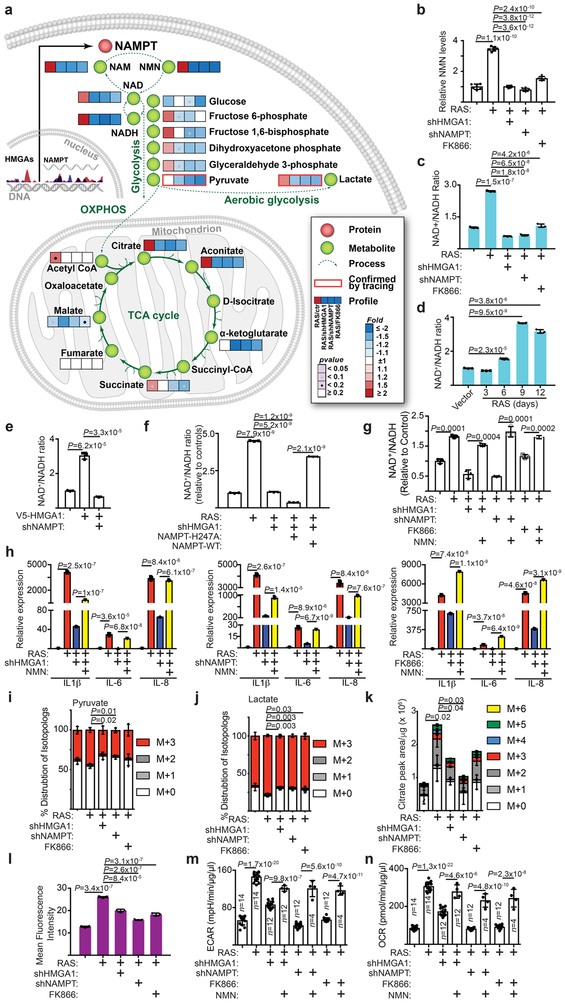
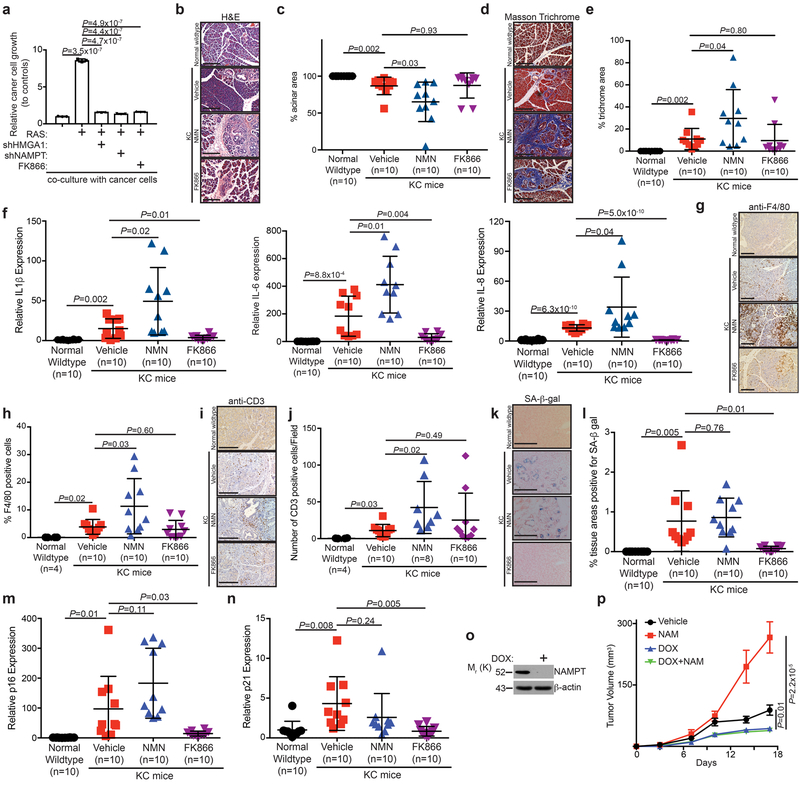
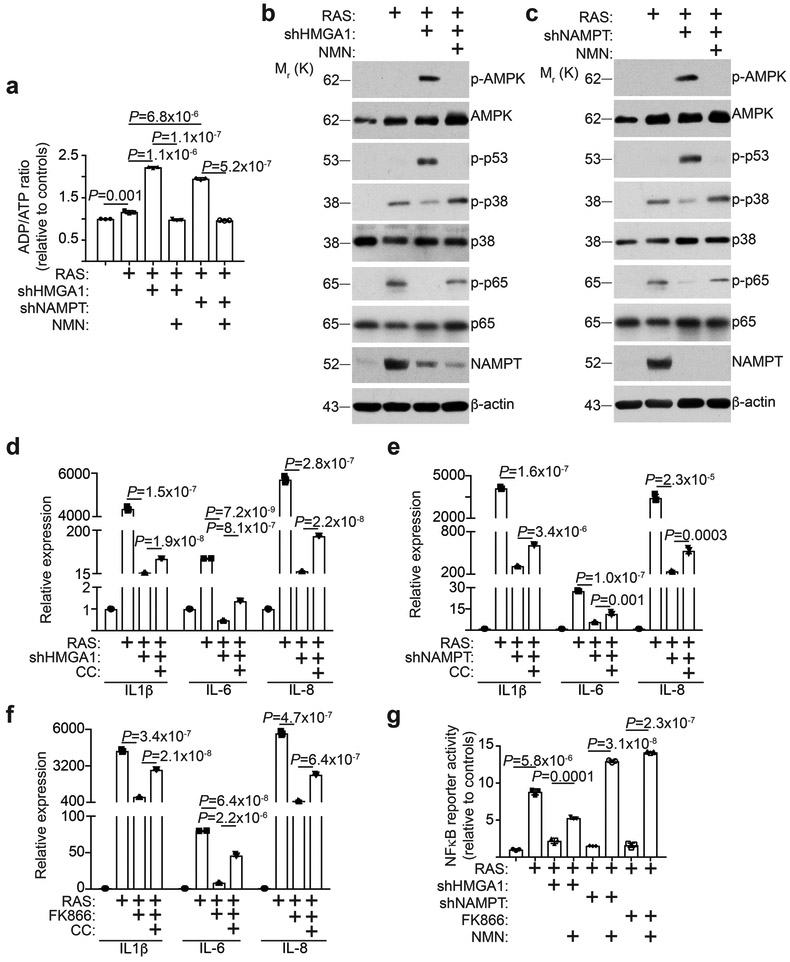
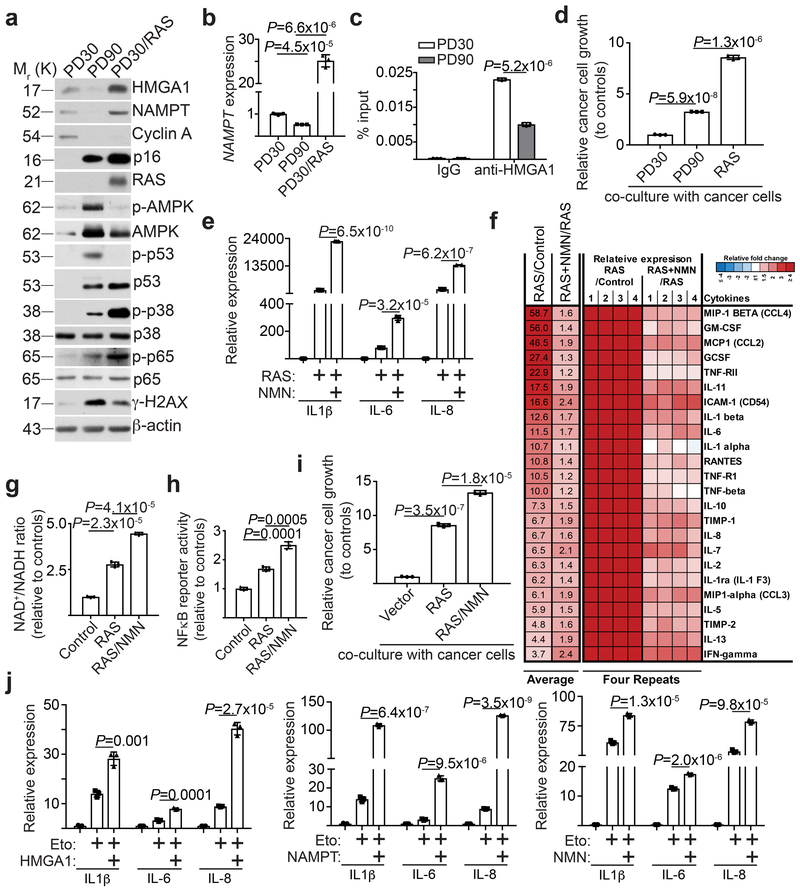
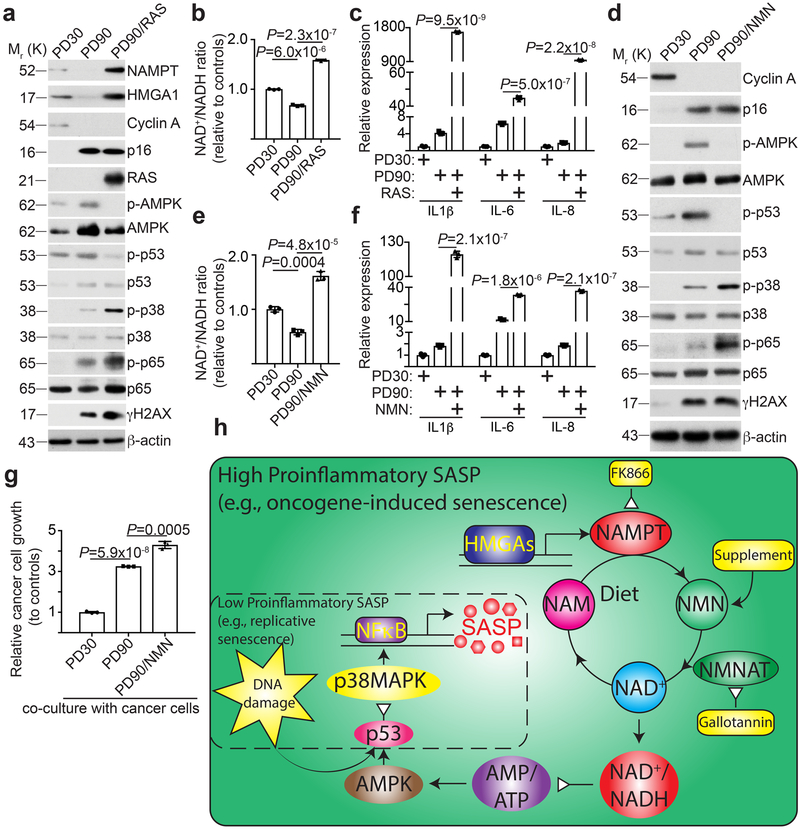
Cellular senescence is a stable growth arrest that is implicated in tissue ageing and cancer. Senescent cells are characterized by an upregulation of proinflammatory cytokines, which is termed the senescence-associated secretory phenotype (SASP). NAD+ metabolism influences both tissue ageing and cancer. However, the role of NAD+ metabolism in regulating the SASP is poorly understood. Here, we show that nicotinamide phosphoribosyltransferase (NAMPT), the rate-limiting enzyme of the NAD+ salvage pathway, governs the proinflammatory SASP independent of senescence-associated growth arrest. NAMPT expression is regulated by high mobility group A (HMGA) proteins during senescence. The HMGA-NAMPT-NAD+ signalling axis promotes the proinflammatory SASP by enhancing glycolysis and mitochondrial respiration. HMGA proteins and NAMPT promote the proinflammatory SASP through NAD+-mediated suppression of AMPK kinase, which suppresses the p53-mediated inhibition of p38 MAPK to enhance NF-κB activity. We conclude that NAD+ metabolism governs the proinflammatory SASP. Given the tumour-promoting effects of the proinflammatory SASP, our results suggest that anti-ageing dietary NAD+ augmentation should be administered with precision.
Figures
References
-
- Hayflick L & Moorhead PS The serial cultivation of human diploid cell strains. Exp Cell Res 25, 585–621 (1961). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK098656/DK/NIDDK NIH HHS/United States
- S10 OD023586/OD/NIH HHS/United States
- P30 CA010815/CA/NCI NIH HHS/United States
- R50 CA221838/CA/NCI NIH HHS/United States
- R00 CA194309/CA/NCI NIH HHS/United States
- F31 CA206387/CA/NCI NIH HHS/United States
- P01 AG031862/AG/NIA NIH HHS/United States
- P50 CA228991/CA/NCI NIH HHS/United States
- R50 CA211199/CA/NCI NIH HHS/United States
- R01 CA202919/CA/NCI NIH HHS/United States
- R01 CA148639/CA/NCI NIH HHS/United States
- R01 CA160331/CA/NCI NIH HHS/United States
- R01 CA131582/CA/NCI NIH HHS/United States
- T32 CA009171/CA/NCI NIH HHS/United States
- R01 CA163377/CA/NCI NIH HHS/United States
- R21 CA155736/CA/NCI NIH HHS/United States
- S10 OD023658/OD/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
